P53 (protein)
|
WikiDoc Resources for P53 (protein) |
|
Articles |
|---|
|
Most recent articles on P53 (protein) Most cited articles on P53 (protein) |
|
Media |
|
Powerpoint slides on P53 (protein) |
|
Evidence Based Medicine |
|
Clinical Trials |
|
Ongoing Trials on P53 (protein) at Clinical Trials.gov Trial results on P53 (protein) Clinical Trials on P53 (protein) at Google
|
|
Guidelines / Policies / Govt |
|
US National Guidelines Clearinghouse on P53 (protein) NICE Guidance on P53 (protein)
|
|
Books |
|
News |
|
Commentary |
|
Definitions |
|
Patient Resources / Community |
|
Patient resources on P53 (protein) Discussion groups on P53 (protein) Patient Handouts on P53 (protein) Directions to Hospitals Treating P53 (protein) Risk calculators and risk factors for P53 (protein)
|
|
Healthcare Provider Resources |
|
Causes & Risk Factors for P53 (protein) |
|
Continuing Medical Education (CME) |
|
International |
|
|
|
Business |
|
Experimental / Informatics |
Editor-In-Chief: C. Michael Gibson, M.S., M.D. [1]
Overview
p53, also known as protein 53 (TP53), is a transcription factor that regulates the cell cycle and hence functions as a tumor suppressor. It is important in multicellular organisms as it helps to suppress cancer. p53 has been described as "the guardian of the genome", "the guardian angel gene", or the "master watchman", referring to its role in conserving stability by preventing genome mutation.[1]
The name p53 is in reference to its apparent molecular mass: it runs as a 53-kilodalton (kDa) protein on SDS-PAGE. But different ways of measuring molecular mass can produce different results. Based on calculations from its amino acid residues, p53's mass is actually only 43.7 kilodaltons. This difference is due to the high number of amino-acid proline residues in the p53 protein which slow p53's migration on SDS-PAGE, thus making it appear larger. This observation is also seen in p53 from other species including rodents, frogs, and fish.
Names
- Official protein name: Cellular tumor antigen p53
Synonyms:
- Tumor suppressor p53
- Phosphoprotein p53
- Antigen NY-CO-13
Gene
The human gene that encodes for p53 is TP53. The gene is named TP53 after the protein it codes for (TP53 is another name for p53). Italics are used to distinguish the TP53 gene name from the TP53 protein name. The gene is located on the human chromosome 17 (17p13.1).
The location has also been mapped on other model animals:
Structure
Human p53 is 393 amino acids long and has five domains:
- An N-terminal transcription-activation domain (TAD), which activates transcription factors : residues 1-42
- A Proline rich domain important for the apoptotic activity of p53 : residues 80-94
- A central DNA-binding core domain (DBD). Contains one zinc atom and several Arginine Amino Acid : Residues 100-300.
- A homo-oligomerisation domain (OD) : residues 307-.355 Tetramerization is essential for the activity of p53 in vivo.
- A C-terminal involved in downregulation of DNA binding of the central domain : residues 356-393.
Mutations that deactivate p53 in cancer usually occur in the DBD. Most of these mutations destroy the ability of the protein to bind to its target DNA sequences, and thus prevents transcriptional activation of these genes. As such, mutations in the DBD are recessive loss-of-function mutations. Molecules of p53 with mutations in the OD dimerise with wild-type p53, and prevent them from activating transcription. Therefore OD mutations have a dominant negative effect on the function of p53.
Wild-type p53 is a labile protein, comprising folded and unstructured regions which function in a synergistic manner.[2]
Functional significance
p53 has many anti-cancer mechanisms:
- It can activate DNA repair proteins when DNA has sustained damage.
- It can also hold the cell cycle at the G1/S regulation point on DNA damage recognition (if it holds the cell here for long enough, the DNA repair proteins will have time to fix the damage and the cell will be allowed to continue the cell cycle.)
- It can initiate apoptosis, the programmed cell death, if the DNA damage proves to be irreparable.

p53 is central to many of the cell's anti-cancer mechanisms. It can induce growth arrest, apoptosis and cell senescence. In normal cells p53 is usually inactive, bound to the protein MDM2 (HDM2 in humans), which prevents its action and promotes its degradation by acting as ubiquitin ligase. Active p53 is induced after the effects of various cancer-causing agents such as UV radiation, oncogenes and some DNA-damaging drugs. DNA damage is sensed by 'checkpoints' in a cell's cycle, and causes proteins such as ATM, CHK1 and CHK2 to phosphorylate p53 at sites that are close to or within the MDM2-binding region and p300-binding region of the protein. Oncogenes also stimulate p53 activation, mediated by the protein p14ARF. Some oncogenes can also stimulate the transcription of proteins which bind to MDM2 and inhibit its activity. Once activated p53 activates expression of several genes including one encoding for p21. p21 binds to the G1-S/CDK and S/CDK complexes (molecules important for the G1/S transition in the cell cycle) inhibiting their activity. p53 has many anticancer mechanisms, and plays a role in apoptosis, genetic stability, and inhibition of angiogenesis.
Recent research has also linked the p53 and RB1 pathways, via p14ARF, raising the possibility that the pathways may regulate each other.[3]
Research published in 2007 showed when p53 expression is stimulated by sunlight, it begins the chain of events leading to tanning.[4]
Regulation of p53 activity
p53 becomes activated in response to a myriad of stress types, which include but is not limited to DNA damage (induced by either UV, IR or chemical agents,such as hydrogen peroxide), oxidative stress, osmotic shock, ribonucleotide depletion and deregulated oncogene expression. This activation is marked by two major events. Firstly, the half-life of the p53 protein is increased drastically, leading to a quick accumulation of p53 in stressed cells. Secondly, a conformational change forces p53 to take on an active role as a transcription regulator in these cells. The critical event leading to the activation of p53 is the phosphorylation of its N-Terminal domain. The N-Terminal transcriptional activation domain contains a large number of phosphorylation sites and can be considered as the primary target for protein kinases transducing stress signals.
The protein kinases which are known to target this transcriptional activation domain of p53, can roughly be divided into two groups. A first group of protein kinases belongs to the MAPK family (JNK1-3, ERK1-2, p38 MAPK), which is known to respond to several types of stress, such as membrane damage, oxidative stress, osmotic shock, heat shock, etc... A second group of protein kinases (ATR, ATM, Chk1, Chk2, DNA-PK, CAK) is implicated in the genome integrity checkpoint, a molecular cascade that detects and responds to several forms of DNA damage caused by genotoxic stress.
In unstressed cells, p53 levels are kept low through a continuous degradation of p53. A protein called Mdm2 binds to p53 and transports it from the nucleus to the cytosol where it becomes degraded by the proteasome. Phosphorylation of the N-terminal end of p53 by the above mentioned protein kinases disrupts Mdm2-binding. Other proteins, such as Pin1, are then recruited to p53 and induce a conformational change in p53 which prevents Mdm2-binding even more. Trancriptional coactivators, like p300 or PCAF, then acetylate the carboxy-terminal end of p53, exposing the DNA binding domain of p53, allowing it to activate or repress specific genes.
Role in disease
If the TP53 gene is damaged, tumor suppression is severely reduced. People who inherit only one functional copy of the TP53 gene will most likely develop tumors in early adulthood, a disease known as Li-Fraumeni syndrome. The TP53 gene can also be damaged in cells by mutagens (chemicals, radiation or viruses), increasing the likelihood that the cell will begin uncontrolled division. More than 50 percent of human tumors contain a mutation or deletion of the TP53 gene. Increasing the amount of p53, which may initially seem a good way to treat tumors or prevent them from spreading, is in actuality not a usable method of treatment, since it can cause premature aging.[5] However, restoring endogenous p53 function holds a lot of promise.[6]
Certain pathogens can also affect the p53 protein that the TP53 gene expresses. One such example, the Human papillomavirus (HPV), encodes a protein, E6, which binds the p53 protein and inactivates it. This, in synergy with the inactivation of another cell cycle regulator, p105RB, allows for repeated cell division manifestested in the clinical disease of warts.
In healthy humans, the p53 protein is continually produced and degraded in the cell. The degradation of the p53 protein is, as mentioned, associated with MDM2 binding. In a negative feedback loop MDM2 is itself induced by the p53 protein. However mutant p53 proteins often don't induce MDM2, and are thus able to accumulate at very high concentrations. Worse, mutant p53 protein itself can inhibit normal p53 protein levels.
Originally P53 was thought to be an oncogene because well over half of all cancer cells tested showed high levels of P53 protein. However, these were all mutated forms of P53.
History
p53 was identified in 1979 by Arnold Levine, David Lane, and Lloyd Old, working at Princeton University, Imperial Cancer Research Fund (UK), and Sloan-Kettering Memorial Hospital, respectively. It had been hypothesized to exist before as the target of the SV40 virus, a strain that induced development of tumors. The TP53 gene from the mouse was first cloned by Peter Chumakov of the Moscow Academy of Sciences in 1982,[7] and independently in 1983 by Moshe Oren (Weizmann Institute).
It was initially presumed to be an oncogene due to the use of mutated cDNA following purification of tumour cell mRNA. Its character as a tumor suppressor gene was finally revealed in 1989 by Bert Vogelstein working at Johns Hopkins School of Medicine.
Warren Maltzman, of the Waksman Institute of Rutgers University first demonstrated that TP53 was responsive to DNA damage in the form of ultraviolet radiation.[8] In a series of publications in 1991-92, Michael Kastan, Johns Hopkins University, reported that TP53 was a critical part of a signal transduction pathway that helped cells respond to DNA damage.
In 1993, p53 was voted molecule of the year by Science magazine.
Its role in sun tanning was published in March 2007.
References
- ↑ Strachan T, Read AP. (1999). Human Molecular Genetics 2. Ch. 18, Cancer Genetics
- ↑ Bell S, Klein C, Muller L, Hansen S, Buchner J (2002) p53 contains large unstructured regions in its native state. J Mol Biol. 2002 Oct 4; 322(5): 917-27;Template:Entrez Pubmed
- ↑ Bates S, Phillips AC, Clark PA, Stott F, Peters G, Ludwig RL, Vousden KH (1998) p14ARF links the tumour suppressors RB and p53. Nature. 1998 Sep 10; 395(6698): 124-5; Template:Entrez Pubmed
- ↑ "Genome's guardian gets a tan started". New Scientist. March 17, 2007. Retrieved 2007-03-29.
- ↑ Tyner SD, Venkatachalam S, Choi J; et al. (2002). "p53 mutant mice that display early ageing-associated phenotypes". Nature. 415 (6867): 45–53. doi:10.1038/415045a. PMID 11780111.
- ↑ Ventura, A. et al. (2007) Restoration of p53 function leads to tumour regression in vivo. Nature. 2007 Feb 8; 445: 661-665; Template:Entrez Pubmed
- ↑ Chumakov P, Iotsova V, Georgiev G (1982). "[Isolation of a plasmid clone containing the mRNA sequence for mouse nonviral T-antigen]". Dokl Akad Nauk SSSR. 267 (5): 1272–5. PMID 6295732.
- ↑ Maltzman W, Czyzyk L (1984). "UV irradiation stimulates levels of p53 cellular tumor antigen in nontransformed mouse cells". Mol Cell Biol. 4 (9): 1689–94. PMID 6092932.
Additional images
-
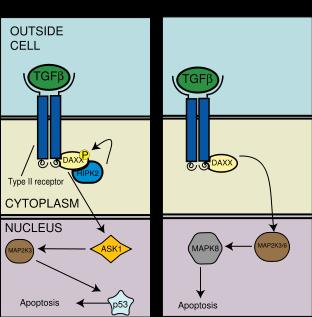 The DAXX Pathway
The DAXX Pathway

{kind=link}
{kind=link}
{kind=link}
External links
- The p53 Protein
- Structure of p53 at genome.ad.jp
- Molecule of the year at hhmi.org
- The p53 Knowledgebase at bii.a-star.edu.sg
- Molecule of the Month at the Protein Data Bank
- the p53 web site
de:P53 it:P53 he:P53 hu:P53 sl:P53 fi:P53 uk:P53 ur:P53 (لحمیہ) Template:WH Template:WikiDoc Sources