Hematopoietic stem cell

Editor-In-Chief: C. Michael Gibson, M.S., M.D. [1]
Overview
Hematopoietic stem cells (HSCs) are stem cells that give rise to all the blood cell types including myeloid (monocytes and macrophages, neutrophils, basophils, eosinophils, erythrocytes, megakaryocytes/platelets, dendritic cells), and lymphoid lineages (T-cells, B-cells, NK-cells). The definition of hematopoietic stem cells has undergone considerable revision in the last two decades. The hematopoietic tissue contains cells with long-term and short-term regeneration capacities and committed multipotent, oligopotent, and unipotent progenitors. Recently, long-term transplantation experiments point toward a clonal diversity model of hematopoietic stem cells. Here, the HSC compartment consists of a fixed number of different types of HSC, each with epigenetically-preprogrammed behavior. This contradicts older models of HSC behavior, which postulated a single type of HSC that can be continuously molded into different subtypes of HSCs. HSCs constitute 1:10.000 of cells in myeloid tissue.

Source
HSCs are found in the bone marrow of adults, which includes femurs, hip, ribs, sternum, and other bones. Cells can be obtained directly by removal from the hip using a needle and syringe, or from the blood following pre-treatment with cytokines, such as G-CSF (granulocyte colony-stimulating factors), that induce cells to be released from the bone marrow compartment. Other sources for clinical and scientific use include umbilical cord blood, placenta, molilized peripheral blood. For experimental purposes, fetal liver, fetal spleen, and AGM (Aorta-gonad-mesonephros) of animals are also useful sources of HSCs.
Functional Characteristics
Multipotency and self-renewal
As stem cells, they are defined by their ability to form multiple cell types (multipotency) and their ability to self-renew.
It is known that a small number of HSCs can expand to generate a very large number of progeny HSCs. This phenomenon is used in bone marrow transplant when a small number of HSCs reconstitute the hematopoietic system. This indicates that, at least during bone marrow transplant, symmetrical cell divisions that give two progeny HSCs must occur, as expansion in HSC numbers seen during bone marrow transplant cannot occur in any other way.
Stem cell self-renewal is thought to occur in the stem cell niche in the bone marrow, and it is reasonable to assume that key signals present in this niche will be important in self-renewal. There is much interest in the environmental and molecular requirements for HSC self-renewal, as understanding the ability of HSC to replenish themselves will eventually allow the generation of expanded populations of HSC ex vivo that can be used therapeutically.
Lineage-Bias
Using limiting dilution strategies combined with other streamlined experimental and statistical methods for examining HSCs at the clonal level, it was shown that HSCs fall into three distinct clusters. These are quantitatively defined by the ratio ρ of lymphoid to myeloid cells that HSC generate upon differentiation (which makes ρ a peripheral predictor for the clonal association of a reconstituted hematopoietic system). Balanced HSCs repopulate peripheral white blood cells in the same ratio of myeloid to lymphoid cells as seen in unmanipulated mice (on average about 15% myeloid and 85% lymphoid cells, or 3≤ρ≤10). Myeloid-biased (My-bi) HSC give rise to too few lymphocytes resulting in ratios 0<ρ<3, whereas lymphoid-biased (Ly-bi) HSC generate too few myeloid cells, which results in lymphoid-to-myeloid ratios of 10<ρ<oo. All three types are normal HSC in that they have self-renewal capacity and can regenerate all hematopietic lineages (pluripotency). What is striking to note is that the lineage-bias is preserved through multiple rounds of serial transplantation: Balanced HSC self-renew to give rise to daughter HSC that are also balanced, My-bi HSC give rise to My-bi daughter HSC, and Ly-bi produce Ly-bi daughter HSC. There is no precursor-progeny relationship between the three types of HSC and they do not represent stages of differentiation. Rather, these are three classes of HSC, each with an epigenetically-fixed differentiation program.
Functional Assays
- Cobble stone area-forming Cell (CAFC) assay: This is a cell culture based empirical assay. When plated onto a confluent culture of stromal feeder layer, a fraction of HSCs creep between the gaps (even though the stromal cells are touching each other) and eventually settle between the stromal cells and the substratum (here the dish surface) or trapped in the cellular processes between the stromal cells. Emperipolesis is the in vivo phenomenon in which one cell is completely engulfed into another (e.g., thymocytes into thymic nurse cells); on the other hand, when in vitro, lymphoid lineage cells creep beneath nurse-like cells, the process is called pseudoemperipolesis. This similar phenomeonon is more commonly known in HSC field by the cell culture terminology cobble stone area-forming cells (CAFC), which means areas of cluster of cells that look dull cobblestone-like under phase contrast microscopy, compared to the other HSCs, which are refractile. This happens because the cells that are floating loosely on top of the stromal cells are spherical and thus refractile. However, the cells that creep beneath the stromal cells are flattened and thus not refractile. The mechanism of pseudoemperipolesis is only recently coming to light. It may be mediated by interaction through CXCR4 (CD184) the receptor for CXC Chemokines (e.g., SDF1) and α4β1 integrins.[1].
Mobility
HSCs has a higher potential than other immature blood cells to pass the bone marrow barrier, and thus may travel in the blood from the bone marrow in one bone to another bone. If they settle in the thymus they'll develop into T cells. In the case of fetuses and other extramedullary hematopoiesis HSCs may also settle in the liver or spleen and develop.
This ability is the reason for that HSCs may be harvested directly from the blood.
Physical characteristics
With regard to morphology, hematopoietic stem cells resemble lymphocytes. They are non-adherent, and rounded, with a rounded nucleus and low cytoplasm-to-nucleus ratio. Since PHSC cannot be isolated as a pure population, it is not possible to identify them in a microscope. The above description is based on the morphological characteristics of a heterogeneous population, of which PHSC are a component.
Markers
In reference to phenotype, hematopoeitic stem cells are identified by their small size, lack of lineage (lin) markers, low staining (side population) with vital dyes such as rhodamine 123 (rhodamineDULL, also called rholo) or Hoechst 33342, and presence of various antigenic markers on their surface, many of which belong to the cluster of differentiation series, like: CD34, CD38, CD90, CD133, CD105, CD45 and also c-kit- the receptor for stem cell factor. The hematopoietic stem cells are negative for the markers that are used for detection of lineage commitment, and are, thus, called Lin-; and, during their purification by FACS, a bunch of up to 14 different mature blood-lineage marker, e.g., CD13 & CD33 for myeloid, CD71 for erythroid, CD19 for B cells, CD61 for megakaryocytic, etc. for humans; and, B220 (murine CD45) for B cells, Mac-1 (CD11b/CD18) for monocytes, Gr-1 for Granulocytes, Ter119 for erythroid cells, Il7Ra, CD3, CD4, CD5, CD8 for T cells, etc. for mice) antibodies are used as a mixture to deplete the lin+ cells or late multipotent progenitors (MPP)s.
There are many differences between the human and mice hematopoietic cell markers for the commonly-accepted type of hematopoietic stem cells.[2].
- Mouse HSC : CD34lo/-, SCA-1+ , Thy1.1+/lo, CD38+, C-kit+, lin-
- Human HSC : CD34+, CD59+, Thy1/CD90+,CD38lo/-, C-kit/CD117-/lo, lin-
However, not all stem cells are covered by these combinations that nonetheless have become popular. In fact, even in humans, there are hematopoietic stem cells that are CD34-/CD38-. [2][3]. Also some later studies suggested that earliest stem cells may lack c-kit on the cell surface[4]. For human HSCs use of CD133 was one step ahead as both CD34+ and CD34- HSCs were CD133+.
Traditional purification method used to yield a reasonable purity level of mouse hematopoietic stem cells, in general, requires a large(~10-12) battery of markers, most of which were surrogate markers with little functional significance, and thus partial overlap with the stem cell populations and sometimes other closely-related cells that are not stem cells. Also, some of these markers 9eg Thy1) are not conserved across mouse species, and use of markers like CD34- for HSC purification requires mice to be at least 8 weeks old. Alternative methods that could give rise to similar or better harvest of stem cells is a hot area of research and are presently emerging. One such method uses a signature of SLAM family of cell surface molecules. SLAM (Signaling lymphocyte activation molecule) family is a group of >10 molecules whose genes are mostly located tandemly in a single locus on chromosome 1 (mouse), all belonging to a subset of immunoglobulin gene superfamily, and originally thought to be involved in T-cell stimulation. This family includes CD48, CD150, CD244, etc., CD150 being the founding member, and, thus, also called slamF1 ie SLAM family member 1.
The signature SLAM code for the hemapoietic higherchy are:
- Hematopoietic stem cells (HSC) : CD150+CD48-CD244-
- Multipotent progenitor cells (MPPs) : CD150-CD48-CD244+
- Lineage-restricted progenitor cells (LRPs) : CD150-CD48+CD244+
For HSCs, CD150+CD48- was sufficient instead of CD150+CD48-CD244- because CD48 is a ligand for CD244, and both would be positive only in the activated lineage-restricted progenitors. It seems that this code was more efficient than the more tedious earlier set of the large number of markers, and are also conserved across the mouse strains; however, recent work has shown that this method excludes a large number of HSCs and includes an equally large number of non-stem cells. [5] [6]. CD150+CD48- gave stem cell purity comparable to Thy1loSca-1+lin-c-kit+ in mice.[7]
Irving Weissman's group at Stanford University that was the first to isolate mouse hematopoietic stem cells in 1988, was also the first to work out the markers to distinguish the mouse long-term (LT-HSC) and short-term (ST-HSC) hematopoietic stem cells (self-renew-capable), and the Multipotent progenitors (MPP, low or no self-renew capability — the later the developmental stage of MPP, the lesser the self-renewal ability and the more of some of the markers like CD4 and CD135):
- LT-HSC : CD34-, SCA-1+ , Thy1.1+/lo, C-kit+, lin-, CD135-, Slamf1/CD150+
- ST-HSC : CD34+, SCA-1+ , Thy1.1+/lo, C-kit+, lin-, CD135-, Slamf1/CD150+, Mac-1 (CD11b)lo
- Early MPP : CD34+, SCA-1+ , Thy1.1-, C-kit+, lin-, CD135+, Slamf1/CD150-, Mac-1 (CD11b)lo, CD4lo
- Late MPP : CD34+, SCA-1+ , Thy1.1-, C-kit+, lin-, CD135high, Slamf1/CD150-, Mac-1 (CD11b)lo, CD4lo
Nomenclature of hematopoietic colonies and lineages
Between 1948 and 1950, the Committee for Clarification of the Nomenclature of Cells and Diseases of the Blood and Blood-forming Organs issued reports on the nomenclature of blood cells.[8][9] An overview of the terminology is shown below, from earliest to final stage of development:
- [root]blast
- pro[root]cyte
- [root]cyte
- meta[root]cyte
- mature cell name
The root for CFU-E is "rubri", for CFU-GM is "granulo" or "myelo" and "mono", for CFU-L is "lympho" and for CFU-Me is "megakaryo". According to this terminology, the stages of red blood cell formation would be: rubriblast, prorubricyte, rubricyte, metarubricyte, and erythrocyte. However, the following nomenclature seems to be, at present, the most prevalent:
| Committee | "lympho" | "rubri" | "granulo" or "myelo" | "mono" | "megakaryo" |
| Lineage | Lymphoid | Myeloid | Myeloid | Myeloid | Myeloid |
| CFU | CFU-L | CFU-E | CFU-GM | CFU-GM | CFU-Me |
| Process | lymphocytopoiesis | erythropoiesis | granulocytopoiesis | monocytopoiesis | thrombocytopoiesis |
| [root]blast | Lymphoblast | Proerythroblast | Myeloblast | Monoblast | Megakaryoblast |
| pro[root]cyte | Prolymphocyte | Polychromatophilic erythrocyte | Promyelocyte | Promonocyte | Promegakaryocyte |
| [root]cyte | - | Normoblast | Eosino/neutro/basophilic myelocyte | Megakaryocyte | |
| meta[root]cyte | Large lymphocyte | Reticulocyte | Eosinophilic/neutrophilic/basophilic metamyelocyte, Eosinophilic/neutrophilic/basophilic band cell | Early monocyte | - |
| mature cell name | Small lymphocyte | Erythrocyte | granulocytes (Eosino/neutro/basophil) | Monocyte | thrombocytes (Platelets) |
Osteoclasts also arise from haemopoietic cells of the monocyte/neutrophil lineage, specifically CFU-GM.
Colony-forming units
There are various kinds of colony-forming units:
- Colony-forming unit lymphocyte (CFU-L)
- Colony-forming unit erythrocyte (CFU-E)
- Colony-forming unit granulo-monocyte (CFU-GM)
- Colony-forming unit megakaryocyte (CFU-Me)
- Colony-forming unit Basophil (CFU-B)
- Colony-forming unit Eosinophil (CFU-Eo)
The above CFUs are based on the lineage. Another CFU, the colony-forming unit–spleen (CFU–S) was the basis of an in vivo clonal colony formation, which depends on the ability of infused bone marrow cells to give rise to clones of maturing hematopoietic cells in the spleens of irradiated mice after 8 to 12 days. It was used extensively in early studies, but is now considered to measure more mature progenitor or Transit Amplifying Cells rather than stem cells.
References
- ↑ Burger JA, Spoo A, Dwenger A, Burger M, Behringer D. CXCR4 chemokine receptors (CD184) and alpha4beta1 integrins mediate spontaneous migration of human CD34+ progenitors and acute myeloid leukaemia cells beneath marrow stromal cells (pseudoemperipolesis). Br J Haematol. 2003 Aug;122(4):579-89. PMID: 12899713
- ↑ Bhatia, M., D. Bonnet, B. Murdoch, O.I. Gan and J.E. Dick, A newly-discovered class of human hematopoietic cells with SCID-repopulating activity, 4(9), 1038, 1998.
- ↑ Guo, Yalin , Lubbert, Michael , Engelhardt, Monika CD34- Hematopoietic Stem Cells: Current Concepts and Controversies Stem Cells 2003; 21: 15-20; First published online ; doi:10.1634/stemcells.21-1-15
- ↑ H. Doi et al. (1997) Proc. Natl. Acad. Sci. USA 94, 2513–2517
- ↑ David C Weksberg, Stuart M Chambers, Nathan C Boles, and Margaret A Goodell CD150 negative Side Population cells represent a functionally-distinct population of long-term hematopoietic stem cells. Blood 2007 : blood-2007-09-115006v1
- ↑ Gary Van Zant Stem cell markers: less is more! Blood 107: 855-856.
- ↑ Kiel et al, Cell, Vol. 121, 1109–1121, July 1, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.cell.2005.05.026
- ↑ "First report of the Committee for Clarification of the Nomenclature of Cells and Diseases of the Blood and Blood-forming Organs". Amer J Clin Pathol. 18: 443–450. 1948.
- ↑ "Third, fourth and fifth reports of the committee for clarification of the nomenclature of cells and diseases of the blood and blood-forming organs". Am J Clin Pathol. 20 (6): 562–79. 1950. PMID 15432355.
Additional images
-
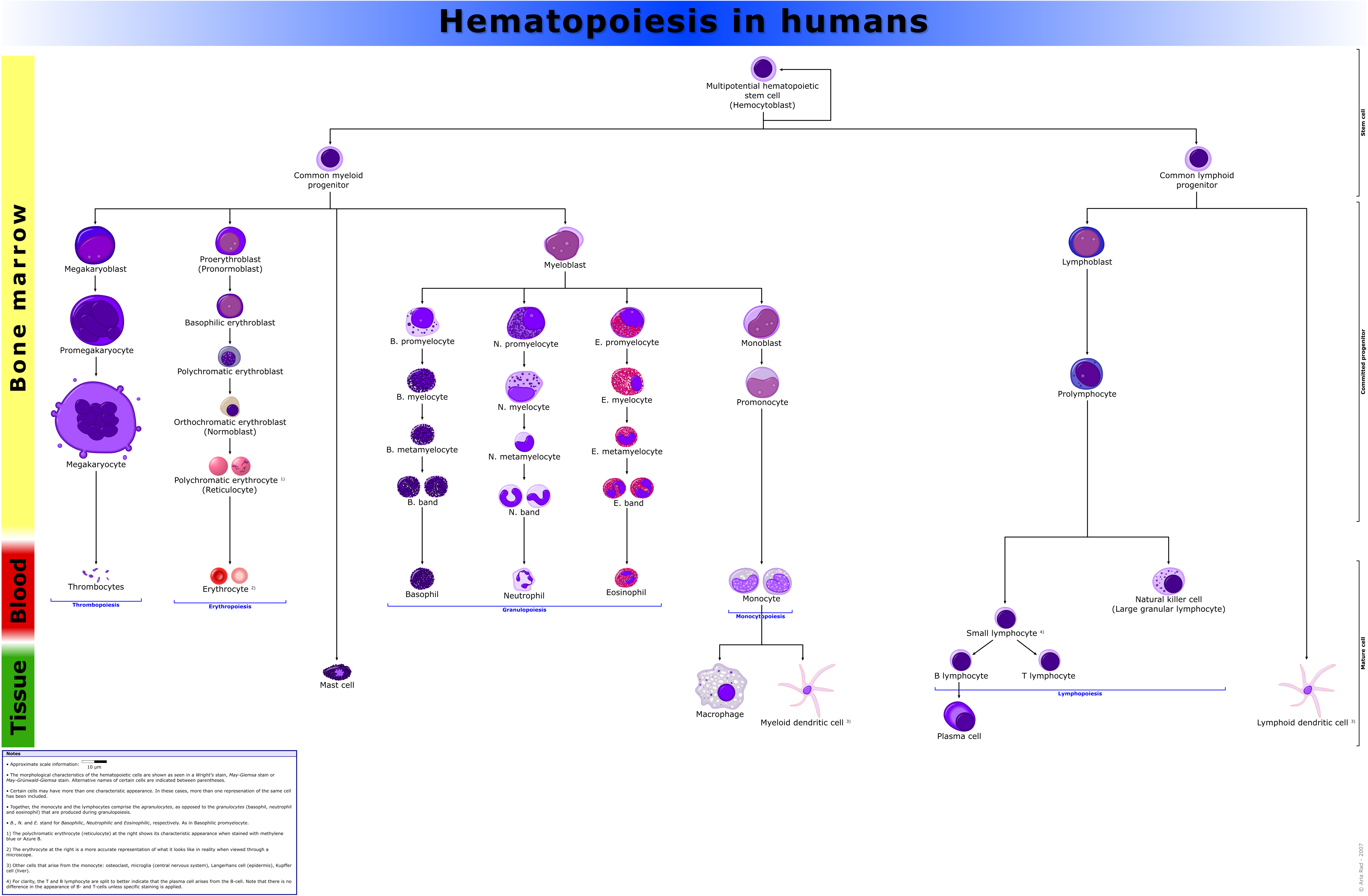 Hematopoiesis
Hematopoiesis
_diagram.png)
See also
External links
- Hematopoietic+stem+cells at the US National Library of Medicine Medical Subject Headings (MeSH)
- Template:EMedicineDictionary