Substantia nigra
The substantia nigra (Latin for "black substance", Sömmering) or locus niger is a heterogeneous portion of the midbrain, separating the pes (foot) from the tegmentum (covering), and a major element of the basal ganglia system. It consists of two strongly contrasted ensembles, the pars compacta and adjacent dopaminergic groups, and another ensemble made up of the pars reticulata and the pars lateralis. The latter two, along with the pallidal nuclei, are elements of the core of the basal ganglia. Although intricate and interconnected, the two ensembles must imperatively be distinguished.
The substantia nigra compacta and surrounding is responsible for dopamine production in the brain, and therefore plays a vital role in reward and addiction.
Pars compacta and adjacent dopaminergic groups
Anatomy
The pars compacta contains neurons which, in humans, are coloured black by the pigment neuromelanin that increases with age. This pigmentation is visible as a distinctive black stripe in brain sections and is the origin of the name given to this area. The neurons have particularly long and thick dendrites (François et al.). The ventral dendrites, particularly, go down deeply in the pars reticulata. Other similar neurons are more sparsely distributed in the mesencephalon and constitute "groups" with no clear borders, although continuous to the pars compacta, in a prerubral position. These have been given in early works in rats with not much respect for the anatomical subdivions the name of "area A8" and "A10". The pars compacta itself ("A9") is usually subdivided into a ventral and a dorsal tier, the last being calbindin positive.[1] The ventral tier is considered as A9v. The dorsal tier A9d is linked to an ensemble comprising also A8 and A10,[2] A8, A9d and A10 representing 28% of dopaminergic neurons. The long dendrites of compacta neurons receive striatal information. This cannot be the case for the more posterior groups that are located outside the striato-pallidonigral bundle territory. Neurons of the pars compacta receive inhibiting signals from the collateral axons from the neurons of the pars reticulata.[3] All these neurons send their axons along the nigrostriatal pathway to the striatum where they release the neurotransmitter dopamine. There is an organization in which dopaminergic neurons of the fringes (the lowest) go to the sensorimotor striatum and the highest to the associative striatum. Dopaminergic axons also innervate other elements of the basal ganglia system including the lateral and medial pallidum,[4] substantia nigra pars reticulata, and the subthalamic nucleus.[5]
Function
The function of the dopamine neurons in the substantia nigra pars compacta is complex. Contrary to what was thought initially it is not directly linked to movements. "Dopamine neurons are activated by novel, unexpected stimuli, by primary rewards in the absence of predictive stimuli and during learning".[6] Dopamine neurons are thought to be involved in learning to predict which behaviours will lead to a reward (for example food or sex). In particular, it is suggested that dopamine neurons fire when a reward is greater than that previously expected; a key component of many reinforcement learning models. This signal can then be used to update the expected value of that action. Many drugs of abuse, such as cocaine, mimic this reward response—providing an explanation for their addictive nature.
Pathology
Degeneration of pigmented neurons in this region is the principal pathology that underlies Parkinson's disease. In a few people, the cause of Parkinson's disease is genetic, but in most cases, the reason for the death of these dopamine neurons is unknown. Parkinsonism can also be produced by viral infections such as encephalitis or a number of toxins, such as MPTP, an industrial toxin which can be mistakenly produced during synthesis of the meperidine analog MPPP. Many such toxins appear to work by producing reactive oxygen species. Binding to neuromelanin by means of charge transfer complexes may concentrate radical-generating toxins in the substantia nigra.
Pathological changes to the dopaminergic neurons of the pars compacta are also thought to be involved in schizophrenia (see the dopamine hypothesis of schizophrenia) and psychomotor retardation sometimes seen in clinical depression.
Pars reticulata and lateralis
Anatomy
Neurons in the pars reticulata and lateralis are much less densely packed than those in the compacta (they were sometimes named pars diffusa). They are smaller and thinner than the dopaminergic neurons and conversely identical and morphologically similar to the pallidal neurons (see primate basal ganglia). Their dendrites as well as the pallidal are preferentially perpendicular to the striatal afferents.[7] The massive striatal afferences corresponds to the medial end of the striato-pallidonigral bundle. Nigral neurons have the same peculiar synaptology with the striatal axonal endings. They make connections with the dopamine neurons of the pars compacta whose long dendrites plunge deeply in the pars reticulata. The neurons of the pars reticulata and lateralis produce the neurotransmitter gamma-aminobutyric acid (GABA). The neurons of the pars reticulata through the nigrothalamic bundle send axons to a particular part of the motor thalamus. The nigral territory corresponds to the nucleus ventralis anterior VA (different from the pallidal VO). VA is the origin of one output of the basal ganglia system. It sends axons to the frontal and oculomotor cortex. In addition the pars reticulata sends neurons to the pars parafascicularis of the central region of the thalamus and to the pedunculopontine complex). The particularity of the pars lateralis is to send its axons to the superior colliculus,[8] which is a too minimized output of the basal ganglia system.
Function
The neurons of the pars reticulata are fast-spiking pacemakers, generating action potentials in the absence of synaptic input.[9] In primates they discharge at a mean rate of 68Hz in contrast to dopaminergic neurons (below 8Hz).[10] They receive abundant afferrences from the striatum (mainly from the associative striatum) with the same very peculiar synaptology as the pallidum. It receives axons from the subthalamic nucleus and a dopaminergic innervation from the dopaminergic ensemble.
The pars reticulata is one of the two primary output nuclei of the basal ganglia system to the motor thalamus (the other output is the internal segment of the globus pallidus).The nigral neurons have their own territory distinct from the cerebellar and the pallidal in the nucleus ventralis anterior VA. This sends axons to the frontal and oculomotor cortex. Hikosaka and Wurtz[11] devoted four papers to "the visual and oculomotor functions of the monkey substantia nigra pars reticulata". This is largely involved in orientation and the control of eye movements in stabilisation of gaze and in saccades.
Pathology
The function of the neurons of the pars reticulata is profoundly changed in parkinsonism and epilepsy. These changes are thought to be mostly secondary to pathology elsewhere in the brain, but may be crucial to understanding the generation of the symptoms of these disorders.
References
- ↑ Francois, C.; Yelnik, J.; Tande, D.; Agid, Y. & Hirsch, E.C. "Dopaminergic cell group A8 in the monkey: anatomical organization and projections to the striatum". Journal of Comparative Neurology. 414 (3): 334–347. PMID 10516600.
- ↑ Feigenbaum Langer, L.; Jimenez-Castellanos, J. & Graybiel, A.M. (1991). "The substantia nigra and its relations with the striatum in the monkey". Progress in Brain Research. 87: 81–99. PMID 1678193.
- ↑ Hajos, M. & Greenfield, S.A. (1994). "Synaptic connections between pars compacta and pars reticulata neurones: electrophysiological evidence for functional modules within the substantia nigra". Brain Research. 660 (2): 216–224. PMID 7820690.
- ↑ Lavoie, B., Smith, Y., Parent, A. (1989). "Dopaminergic innervation of the basal ganglia in the squirrel monkey as revealed by tyrosine hydroxylase immunohistochemistry". 289 (1): 36–52. PMID 2572613.
- ↑ Cragg S.J.; Baufreton J.; Xue Y.; Bolam J.P.; & Bevan M.D. (2004). "Synaptic release of dopamine in the subthalamic nucleus". European Journal of Neuroscience. 20 (7): 1788–1802. PMID 15380000.
- ↑ Schultz, W. (1992). "Activity of dopamine neurons in the behaving primate". Seminar in Neuroscience. 4: 129–138.
- ↑ François, C.; Yelnik, J. & Percheron, G. (1987). "Golgi study of the primate substantia nigra. II. Spatial organization of dendritic arborizations in relation to the cytoarchitectonic boundaries and to the striatonigral bundle". Journal of Comparative Neurology. 265 (4): 473–493. PMID 3123530.
- ↑ François, C.; Percheron, G. & Yelnik, J. (1984). "Localization of nigrostriatal, nigrothalamic and nigrotectal neurons in ventricular coordinates in macaques". Neuroscience. 13 (1): 61–76. PMID 6387531.
- ↑ Atherton, J.F. & Bevan, M.D. (2005). "Ionic mechanisms underlying autonomous action potential generation in the somata and dendrites of GABAergic substantia nigra pars reticulata neurons in vitro". Journal of Neuroscience. 25 (36): 8272–8281. PMID 16148235.
- ↑ Schultz, W. (1986). "Activity of pars reticulata neurons of monkey substantia nigra in relation to motor, sensory and complex events". Journal of Neurophysiology. 55 (4): 660–677. PMID 3701399.
- ↑ Hikosaka, O. & Wurtz, R.H. (1983). "Visual and oculomotor functions of monkey substantia nigra pars reticulata. I to IV". Journal of Neurophysiology. 49 (5): 1230–1301. PMID 6864248, PMID 6864249, PMID 6864250 & PMID 6306173.
Uh huh
See also
Additional images
-
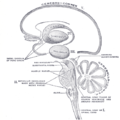 Schematic representation of the chief ganglionic categories (I to V).
Schematic representation of the chief ganglionic categories (I to V). -
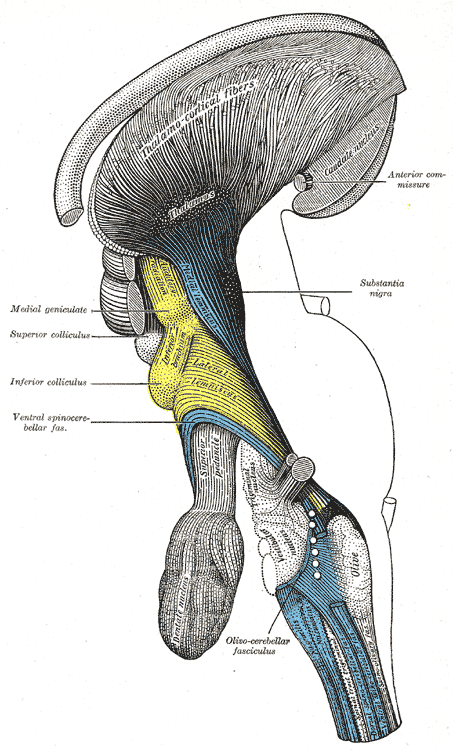 Deep dissection of brain-stem. Lateral view.
Deep dissection of brain-stem. Lateral view. -
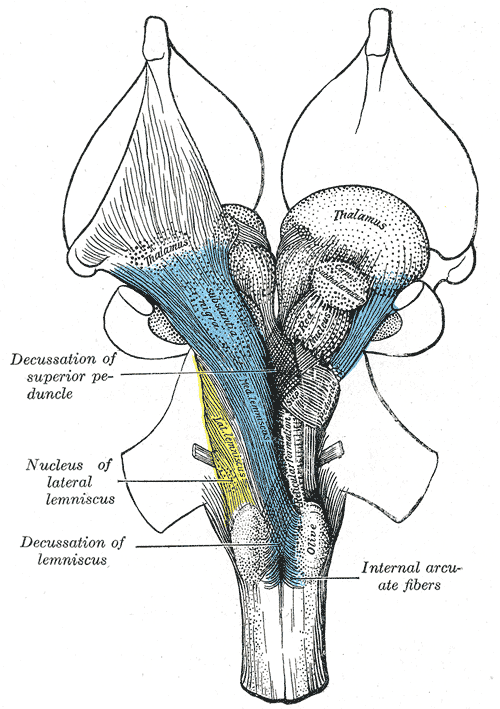 Deep dissection of brain-stem. Ventral view.
Deep dissection of brain-stem. Ventral view. -
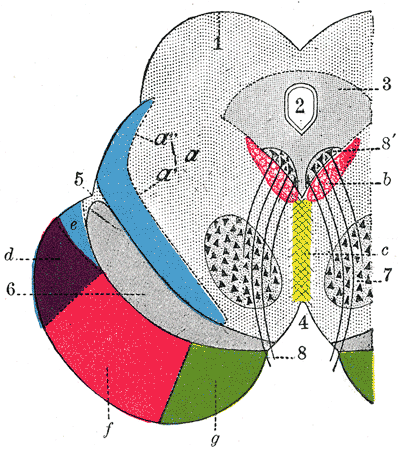 Coronal section through mid-brain.
Coronal section through mid-brain. -
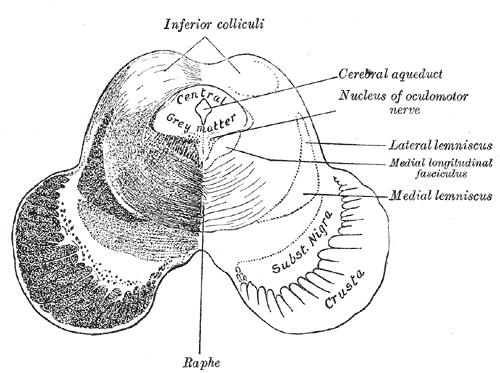 Transverse section of mid-brain at level of inferior colliculi.
Transverse section of mid-brain at level of inferior colliculi. -
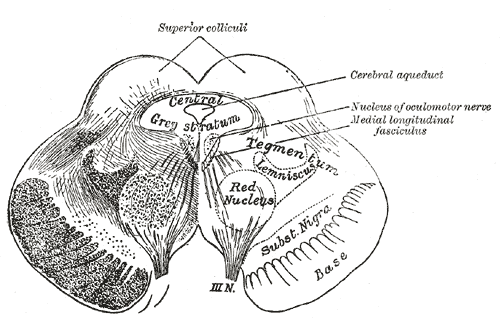 Transverse section of mid-brain at level of superior colliculi.
Transverse section of mid-brain at level of superior colliculi. -
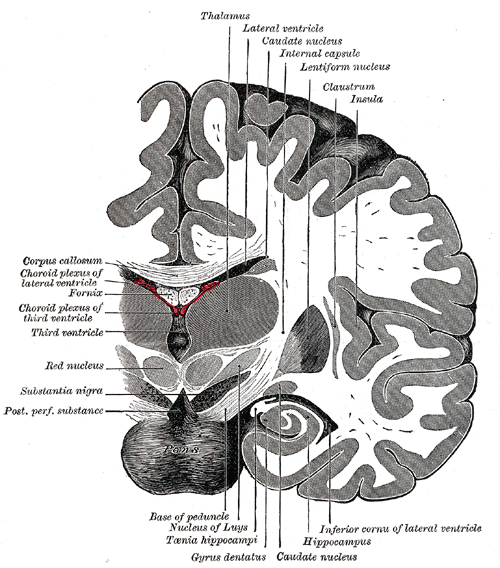 Coronal section of brain immediately in front of pons.
Coronal section of brain immediately in front of pons.







External links
- Template:BrainstemWisconsin
- Template:BrainMaps
- Diagram at uni-tuebingen.de
- Diagram and explanation of cortico-basal pathways
{kind=link}
bg:Субстанция нигра da:Sorte substans de:Substantia nigra it:Substantia nigra nl:Substantia nigra no:Substantia nigra