Cochlear nuclei
Overview
The cochlear nuclei consist of:
- (a) the dorsal cochlear nucleus, corresponding to the tuberculum acusticum on the dorso-lateral surface of the inferior peduncle; and
- (b) the ventral or accessory cochlear nucleus, placed between the two divisions of the nerve, on the ventral aspect of the inferior peduncle.
Anatomy and function
The CN is the first relay station in the auditory system and is located at the dorso-lateral side of the brainstem, spanning the junction of the pons and medulla. Information is brought via the cochlear nerve, also called Cranial nerve VIII, to the CN. The cochlear nucleus can be divided into ventral and dorsal sections (DCN). The ventral part can further be divided into anterior and posterior sections (AVCN and PVCN), which are separated by the incoming auditory nerve fibers. The auditory nerve fibers form a highly organized system of connections according to their peripheral innervation of the cochlea. Axons from the spiral ganglion cells of the lower frequency innervate the ventral portions of the dorsal cochlear nucleus and the ventrolateral portions of the anteroventral cochlear nucleus. In contrast, the axons from the higher frequency organ of corti hair cells project to the dorsal portion of the anteroventral cochlear nucleus and the uppermost dorsal portions of the dorsal cochlear nucleus. The mid frequency projections end up in between the two extremes, in this way the frequency spectrum is preserved.
At the level of the cochlear nuclei, the input from the two ears, for the most part, remains separated. There is evidence, however, that a significant number of neurons in the CN receives input from the contralateral CN. Just as the inner hair cells are arranged according to the best frequency (BF), so is the cochlea nucleus. This so-called tonotopic organization is preserved because only a few inner hair cells synapse on the dendrites of a nerve cell in the spiral ganglion, and the axon from that nerve cell synapes on only a very few dendrites in the cochlear nuclues.
Each cochlear nucleus has two parts, dorsal (DCN) and ventral (VCN). The Cochlear Nucleus receives input from each spiral ganglion, and also receives input from other parts of the brain. How the inputs from other areas of the brain affect hearing is unknown.
Location and projections from the CN
There are three major projections from the cochlear nuclei. Through the medulla, one projection bifurcates, and projects to the contralateral the superior olivary complex (SOC) via the trapezoid body, whilst the other half shoots to the ipsilateral SOC. This projection is called the ventral acoustic stria (or, more commonly, the trapezoid body). Another projection, called the dorsal acoustic stria (DAS, also known as the stria of von Monakow), rises above the medulla into the pons where it hits the nucleus of the lateral lemniscus along with its kin, the intermediate acoustic stria (IAS, also known as the stria of Held). The IAS decussates across the medulla, before joining the ascending fibers in the contralateral lateral lemniscus. The lateral lemniscus contains cells of the nuclei of the lateral lemniscus, and in turn projects to the inferior colliculus. The inferior colliculus receives direct, monosynaptic projections from the superior olivary complex the contralateral dorsal acoustic stria, some classes of stellate neurons of the VCN, as well as from the different nuclei of the lateral lemniscus. All of these inputs terminate in the inferior colliculus, although there are a few small projections that bypass the inferior colliculus and project to the medial geniculate, or other forebrain structures. The principal projection of the inferior colliculus is to the medial geniculate body in the thalamus. The medial geniculate then projects to the auditory cortex, in the superior temporal gyrus.
Cell types
There are four types of principal cells found in the cochlear nuclei: Bushy cells, stellate cells, octopus cells, and fusiform cells.
- Bushy cells are found in the anterior ventral cochlear nucleus (AVCN). These can be divided into spherical and globular bushy cells, depending on their appearance, and also their location. Within the AVCN here is an area of large spherical cells; caudal to this are smaller spherical cells, and globular cells. They have a few (1-4) very short dendrites with numerous small branching, which cause it to resemble a “bush”. The bushy cells are only found in the ventral portion of the cochlear nucleus. The bushy cells have specialized electrical properties that allow them to transmit timing information from the auditory nerve to more central areas of the auditory system. Some bushy cells can even improve the precision of the timing information. Bushy cells have responses very similar to those in the auditory nerve. The primary difference is that spontaneous activity is decreased by stimulation by adjacent frequencies, therefore leading to an even sharper tuning curve than seen in auditory nerve cells. These cells are usually innervated only by a selected few axons, which dominate its firing patterns. These afferent axons wrap their terminal branches around the entire soma, creating a large synapse onto the bushy cells. Therefore, a single unit recording of an electrically stimulated bushy neuron characteristically produces exactly one action potential and constitutes the primary response.
- Stellate cells, morphologically, have a radial, star-like dendritic tree, which is where they get their name. They are also called, chopper cells, in reference to their ability to fire a regularly-spaced train of action potentials for the duration of a tonal or noise stimulus. The chopping pattern is intrinsic to the electrical excitability of the stellate cell, and the firing rate depends on the strength of the auditory input more than on the frequency.
- Octopus cells are found in a small region of the Posterior Ventral Cochlear Nucleus (PVCN). The distinguishing features of these cells are their long, thick dendrites that typically emanate from one side of the cell body. Octopus cells produce an "Onset Response" to simple tonal stimuli. That is, they respond only at the onset of a specific frequency or frequency range at higher amplitudes. The octopus cells can fire with some of the highest temporal precision of any neuron in the brain. Electrical stimuli to the auditory nerve has been shown to evoke a graded post synaptic potential in the octopus cells. These EPSP’s are very brief. The octopus cells are thought to be important extracting timing information. It has been reported that these cells can respond to click trains at a rate of 800 Hz.
- Fusiform cells (also known as pyramidal cells) are found in the Dorsal Cochlear Nucleus (DCN). See the separate page concerning the DCN.
Structures
The input of auditory stimulus is through the Auditory Nerve (CN VIII). The auditory stimulus further travels through the AVCN as neural impulses and branches through the following structures:
- Medial superior olive (MSO) via Trapezoid Body (TB) – Ipsilateral and contralateral stimulation for low frequency sounds.
- Lateral superior olive (LSO) directly and via TB – Ipsilateral stimulation for high frequency sounds.
- Medial Nucleus of Trapezoid body (MNTB) – Contralateral stimulation.
- Inferior colliculus – Contralateral stimulation.
- Neurotransmitters: There are four neurotransmitters responsible for transmission of neural impulses, namely, GABA, Norepinephrine, Glutamate, and Acetylcholine.
- Periolivary nuclei (PON) – Ipsilateral and Contralateral stimulation.
- Lateral lemniscus (LL) and Lemniscal Nuclei (LN) – Ipsilateral and Contralateral Stimulation.
- Inferior colliculus – Contralateral stimulation.
See also
References
Additional images
-
 Dissection of brain-stem. Lateral view.
Dissection of brain-stem. Lateral view. -
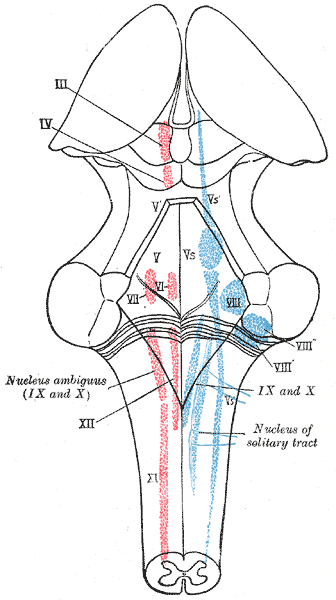 The cranial nerve nuclei schematically represented; dorsal view. Motor nuclei in red; sensory in blue.
The cranial nerve nuclei schematically represented; dorsal view. Motor nuclei in red; sensory in blue. -
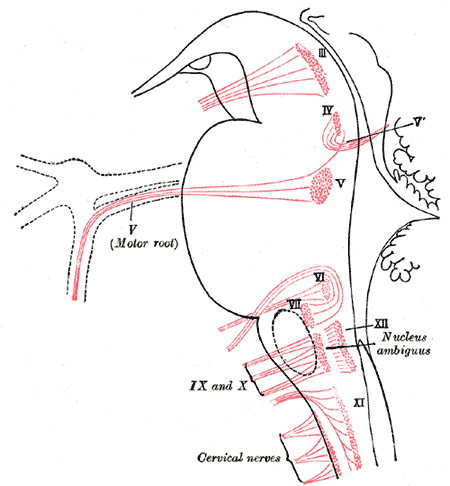 Nuclei of origin of cranial motor nerves schematically represented; lateral view.
Nuclei of origin of cranial motor nerves schematically represented; lateral view. -
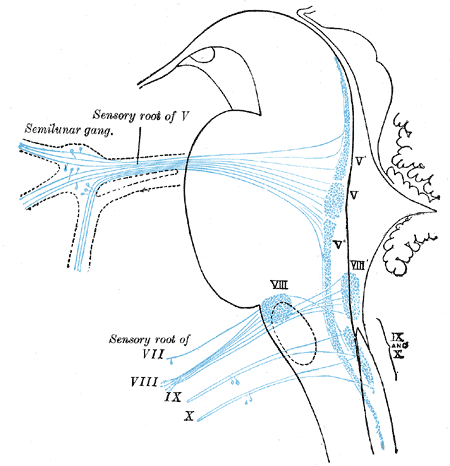 Primary terminal nuclei of the afferent (sensory) cranial nerves schematically represented; lateral view.
Primary terminal nuclei of the afferent (sensory) cranial nerves schematically represented; lateral view. -
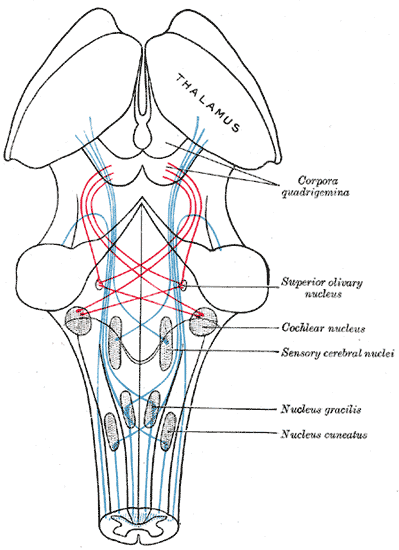 Scheme showing the course of the fibers of the lemniscus; medial lemniscus in blue, lateral in red.
Scheme showing the course of the fibers of the lemniscus; medial lemniscus in blue, lateral in red.





External links
Template:Gray's Template:Auditory system Template:Rhombencephalon