Karyotype

|
WikiDoc Resources for Karyotype |
|
Articles |
|---|
|
Most recent articles on Karyotype |
|
Media |
|
Evidence Based Medicine |
|
Clinical Trials |
|
Ongoing Trials on Karyotype at Clinical Trials.gov Clinical Trials on Karyotype at Google
|
|
Guidelines / Policies / Govt |
|
US National Guidelines Clearinghouse on Karyotype
|
|
Books |
|
News |
|
Commentary |
|
Definitions |
|
Patient Resources / Community |
|
Patient resources on Karyotype Discussion groups on Karyotype Directions to Hospitals Treating Karyotype Risk calculators and risk factors for Karyotype
|
|
Healthcare Provider Resources |
|
Causes & Risk Factors for Karyotype |
|
Continuing Medical Education (CME) |
|
International |
|
|
|
Business |
|
Experimental / Informatics |
Editor-In-Chief: C. Michael Gibson, M.S., M.D. [1]
Overview
A karyotype is the characteristic chromosome complement of an eukaryote species.[1][2] The preparation and study of karyotypes is part of cytogenetics.
In normal diploid organisms, autosomal chromosomes are present in two identical copies. There may, or may not, be sex chromosomes. Polyploid cells have multiple copies of chromosomes and haploid cells have single copies. The study of whole sets of chromosomes is sometimes known as karyology. The chromosomes are depicted (by rearranging a microphotograph) in a standard format known as a karyogram or idiogram: in pairs, ordered by size and position of centromere for chromosomes of the same size.
The study of karyotypes is made possible by staining: usually a suitable dye is applied after cells have been arrested during cell division by a solution of colchicine.[3] Sometimes observations may be made on non-dividing (interphase) cells. The sex of an unborn fetus can be determined by observation of interphase cells (see amniotic centesis and Barr body).
Most (but not all) species have a standard karyotype. The normal human karyotypes contain 22 pairs of autosomal chromosomes and one pair of sex chromosomes. Normal karyotypes for women contain two X chromosomes and are denoted 46,XX; men have both an X and a Y chromosome denoted 46,XY. However, some individuals have other karyotypes with added or missing chromosomes, and in all such cases there are developmental abnormalities as a consequence.
Karyotypes can be used for many purposes. They may be used to study chromosomal aberrations, to study cellular function, to study taxonomic relationships, or to gather information about past evolutionary events.
Observations on karyotypes
Six different characteristics of karyotypes are usually observed and compared:[4]
- differences in absolute sizes of chromosomes. Chromosomes can vary in absolute size by as much as twenty-fold between genera of the same family: Lotus tenuis and Vicia faba (legumes), both have six pairs of chromosomes (n=6) yet V. faba chromosomes are many times larger. This feature probably reflects different amounts of DNA duplication.
- differences in the position of centromeres. This is brought about by translocations.
- differences in relative size of chromosomes can only be caused by segmental interchange of unequal lengths.
- differences in basic number of chromosomes may occur due to successive unequal translocations which finally remove all the essential genetic material from a chromosome, permitting its loss without penalty to the organism (the dislocation hypothesis). Humans have one pair fewer chromosomes than the great apes, but the genes have been mostly translocated (added) to other chromosomes.
- differences in number and position of satellites, which (when they occur) are small bodies attached to a chromosome by a thin thread.
- differences in degree and distribution of heterochromatic regions. Heterochromatin stains darker than euchromatin, indicating tighter packing, and mainly consists of genetically inactive repetitive DNA sequences.
A full account of a karyotype may therefore include the number, type, shape and banding of the chromosomes, as well as other cytogenetic information.
Variation is often found:
- between the two sexes
- between the germ-line and soma (between gametes and the rest of the body)
- between members of a population (chromosome polymorphism)
- geographical variation between races
- mosaics or otherwise abnormal individuals.[5]
Historical note
Levitsky seems to have been the first to define the karyotype as the phenotypic appearance of the somatic chromosomes, in contrast to their genic contents.[6][7] The subsequent history of the concept can be followed in the works of Darlington[8] and White.[9][10]
Investigation into the human karyotype took many years to settle the most basic question: how many chromosomes does a normal diploid human cell contain?[11] In 1912, Hans von Winiwarter reported 47 chromosomes in spermatogonia and 48 in oogonia, concluding an XX/XO sex determination mechanism.[12] Painter in 1922 was not certain whether the diploid number of man was 46 or 48, at first favoring 46.[13] He revised his opinion later from 46 to 48, and he correctly insisted on man having an XX/XY system.[14] Considering their techniques, these results were quite remarkable.
New techniques were needed to definitively solve the problem:
- Using cells in culture
- Pretreating cells in a hypotonic solution, which swells them and spreads the chromosomes
- Arresting mitosis in metaphase by a solution of colchicine
- Squashing the preparation on the slide forcing the chromosomes into a single plane
- Cutting up a photomicrograph and arranging the result into an indisputable karyogram.
It took until the mid 1950s until it became generally accepted that the karyotype of man included only 46 chromosomes.[15][16] Rather interestingly, the great apes have 48 chromosomes.
Diversity and evolution of karyotypes
Although the replication and transcription of DNA is highly standardized in eukaryotes, the same cannot be said for their karyotypes, which are highly variable between species in chromosome number and in detailed organization despite being constructed out of the same macromolecules. In some cases there is even significant variation within species. This variation provides the basis for a range of studies in what might be called evolutionary cytology.
Changes during development
Instead of the usual gene repression, some organisms go in for large-scale elimination of heterochromatin, or other kinds of visible adjustment to the karyotype.
- Chromosome elimination. In some species, as in many sciarid flies, entire chromosomes are eliminated during development.[17]
- Chromatin diminution (founding father: Theodor Boveri). In this process, found in some copepods and roundworms such as Ascaris suum, portions of the chromosomes are cast away in particular cells. This process is a carefully organised genome rearrangement where new telomeres are constructed and certain heterochromatin regions are lost.[18][19] In A. suum, all the somatic cell precursors undergo chromatin diminution.[20]
- X-inactivation. The inactivation of one X chromosome takes place during the early development of mammals (see Barr body and dosage compensation). In placental mammals, the inactivation is random as between the two Xs; but in marsupials it is always the paternal X which is inactivated.
Number of chromosomes in a set
A spectacular example of variability between closely related species is the muntjac, which was investigated by Kurt Benirschke and his colleague Doris Wurster. The diploid number of the Chinese muntjac, Muntiacus reevesi, was found to be 46, all telocentric. When they looked at the karyotype of the closely related Indian muntjac, Muntiacus muntjak, they were astonished to find it had female = 6, male = 7 chromosomes.[21]
- "They simply could not believe what they saw... They kept quiet for two or three years because they thought something was wrong with their tissue culture... But when they obtained a couple more specimens they confirmed [their findings]"[22]
The number of chromosomes in the karyotype between (relatively) unrelated species is hugely variable. The low record is held by the nematode Parascaris univalens, where the haploid n = 1; the high record would be somewhere amongst the ferns, with the Adder's Tongue Fern Ophioglossum ahead with an average of 1262 chromosomes.[23] Top score for animals might be the shortnose sturgeon Acipenser brevirostrum at a mere 372 chromosomes.[24] The existence of supernumerary or B chromosomes means that chromosome number can vary even within one interbreeding population; and aneuploids are another example, though in this case they would not be regarded as normal members of the population.
Ploidy: the number of sets in a karyotype
- Polyploidy, where there are more than two sets of homologous chromosomes in the cells, occurs mainly in plants. It has been of major significance in plant evolution according to Stebbins.[25][26][27][28] The proportion of flowering plants which are polyploid was estimated by Stebbins to be 30-35%, but in grasses the average is much higher, about 70%.[29] Polyploidy in lower plants (ferns, horsetails and psilotales) is also common, and some species of ferns have reached levels of polyploidy far in excess of the highest levels known in flowering plants.
Polyploidy in animals is much less common, but it has been significant in some groups.[30]
- Haplo-diploidy, where one sex is diploid, and the other haploid. It is a common arrangement in the Hymenoptera, and in some other groups.
- Endopolyploidy occurs when in adult differentiated tissues the cells have ceased to divide by mitosis, but the nuclei contain more than the original somatic number of chromosomes.[31] In the endocycle (endomitosis or endoreduplication) chromosomes in a 'resting' nucleus undergo reduplication, the daughter chromosomes separating from each other inside an intact nuclear membrane.[32]
In many instances, endopolyploid nuclei contain tens of thousands of chromosomes (which cannot be exactly counted). The cells do not always contain exact multiples (powers of two), which is why the simple definition 'an increase in the number of chromosome sets caused by replication without cell division' is not quite accurate.
This process (especially studied in insects and some higher plants such as maize) may be a developmental strategy for increasing the productivity of tissues which are highly active in biosynthesis.[33]
The phenomenon occurs sporadically throughout the eukaryote kingdom from protozoa to man; it is diverse and complex, and serves differentiation and morphogenesis in many ways.[34]
- See palaeopolyploidy for the investigation of ancient karyotype duplications.
Species trees
The detailed study of chromosome banding in insects with polytene chromosomes can reveal relationships between closely related species: the classic example is the study of chromosome banding in Hawaiian drosophilids by Hampton Carson.
In about 6,500 square miles, the Hawaiian islands have the most diverse collection of drosophilid flies in the world, living from rainforests to subalpine meadows. These roughly 800 Hawaiian drosophilid species are usually assigned to two genera Drosophila and Scaptomyza in the family Drosophilidae.
The polytene banding of the 'picture wing' group, the best-studied group of Hawaiian drosophilids, enabled Carson to work out the evolutionary tree long before genome analysis was practicable. In a sense, gene arrangements are visible in the banding patterns of each chromosome. Chromosome rearrangements, especially inversions, make it possible to see which species are closely related.
The results are clear. The inversions, when plotted in tree form (and independent of all other information), show a clear "flow" of species from older to newer islands. There are also cases of colonization back to older islands, and skipping of islands, but these are much less frequent. Using K-Ar dating, the present islands date from 0.4 million years ago (mya) (Mauna Kea) to 10mya (Necker). The oldest member of the Hawaiian archipelago still above the sea is Kure Atoll, which can be dated to 30 mya. The archipelago itself (produced by the Pacific plate moving over a hot spot) has existed for far longer, at least into the Cretaceous. Previous islands now beneath the sea (guyots) form the [[Emperor Seamount Chain.[35]
All of the native Drosophila and Scaptomyza species in Hawaii have apparently descended from a single ancestral species that colonized the islands, probably 20 million years ago. The subsequent adaptive radiation was spurred by a lack of competition and a wide variety of niches. Although it would be possible for a single gravid female to colonise an island, it is more likely to have been a group from the same species.[36][37][38][39]
There are other animals and plants on the Hawaiian archipelago which have undergone similar, if less spectacular, adaptive radiations.[40][41]
Overview
Although much is known about karyotypes at the descriptive level, and it is clear that changes in karyotype organization has had effects on the evolutionary course of many species, it is quite unclear what the general significance might be.
- "We have a very poor understanding of the causes of karyotype evolution, despite many careful investigations... the general significance of karyotype evolution is obscure." Maynard Smith.[42]
Depiction of karyotypes
Types of banding
Cytogenetics employs several techniques to visualize different aspects of chromosomes:[43]
- G-banding is obtained with Giemsa stain following digestion of chromosomes with trypsin. It yields a series of lightly and darkly stained bands - the dark regions tend to be heterochromatic, late-replicating and AT rich. The light regions tend to be euchromatic, early-replicating and GC rich. This method will normally produce 300-400 bands in a normal, human genome.
- R-banding is the reverse of G-banding (the R stands for "reverse"). The dark regions are euchromatic (guanine-cytosine rich regions) and the bright regions are heterochromatic (thymine-adenine rich regions).
- C-banding: Giemsa binds to constitutive heterochromatin, so it stains centromeres.
- Q-banding is a fluorescent pattern obtained using quinacrine for staining. The pattern of bands is very similar to that seen in G-banding.
- T-banding: visualize telomeres.
Classic karyotype cytogenetics

In the "classic" (depicted) karyotype, a dye, often Giemsa (G-banding), less frequently Quinacrine, is used to stain bands on the chromosomes. Giemsa is specific for the phosphate groups of DNA. Quinacrine binds to the adenine-thymine-rich regions. Each chromosome has a characteristic banding pattern that helps to identify them; both chromosomes in a pair will have the same banding pattern.
Karyotypes are arranged with the short arm of the chromosome on top, and the long arm on the bottom. Some karyotypes call the short and long arms p and q, respectively. In addition, the differently stained regions and sub-regions are given numerical designations from proximal to distal on the chromosome arms. For example, Cri du chat syndrome involves a deletion on the short arm of chromosome 5. It is written as 46,XX,5p-. The critical region for this syndrome is deletion of 15.2, which is written as 46,XX,del(5)(p15.2).[44]
Spectral karyotype (SKY technique)

Spectral karyotyping is a molecular cytogenetic technique used to simultaneously visualize all the pairs of chromosomes in an organism in different colors. Fluorescently-labeled probes for each chromosome are made by labeling chromosome-specific DNA with different fluorophores. Because there are a limited number of spectrally-distinct fluorophores, a combinatorial labeling method is used to generate many different colors. Spectral differences generated by combinatorial labeling are captured and analyzed by using an interferometer attached to a fluorescence microscope. Image processing software then assigns a pseudo color to each spectrally different combination, allowing the visualization of the individually colored chromosomes.[45]
This technique is used to identify structural chromosome aberrations in cancer cells and other disease conditions when Giemsa banding or other techniques are not accurate enough.
Digital Karyotyping
Digital Karyotyping is a technique used to quantify the DNA copy number on a genomic scale. Short sequences of DNA from specific loci all over the genome are isolated and enumerated. [46]
Chromosome abnormalities
Chromosome abnormalities can be numerical, as in the presence of extra or missing chromosomes, or structural, as in translocations, inversions, large-scale deletions or duplications. Numerical abnormalities, also known as aneuploidy, often occur as a result of nondisjunction during meiosis in the formation of a gamete; trisomies, in which three copies of a chromosome are present instead of the usual two, are common numerical abnormalities. Structural abnormalities often arise from errors in homologous recombination. Both types of abnormalities can occur in gametes and therefore will be present in all cells of an affected person's body, or they can occur during mitosis and give rise to a genetic mosaic individual who has some normal and some abnormal cells.
Chromosomal abnormalities that lead to disease in humans include:
- Turner syndrome results from a single X chromosome (45, X or 45, X0).
- Klinefelter syndrome, the most common male chromosomal disease, otherwise known as 47, XXY is caused by an extra X chromosome.
- Edwards syndrome is caused by trisomy (three copies) of chromosome 18.
- Down syndrome, a common chromosomal disease, is caused by trisomy of chromosome 21.
- Patau syndrome is caused by trisomy of chromosome 13.
- Also documented are trisomy 8, trisomy 9 and trisomy 16, although they generally do not survive to birth.
Some disorders arise from loss of just a piece of one chromosome, including
- Cri du chat (cry of the cat), from a truncated short arm on chromosome 5. The name comes from the babies' distinctive cry, caused by abnormal formation of the larynx.
- 1p36 Deletion syndrome, from the loss of part of the short arm of chromosome 1.
- Angelman syndrome – 50% of cases have a segment of the long arm of chromosome 15 missing.
Chromosomal abnormalities can also occur in cancerous cells of an otherwise genetically normal individual; one well-documented example is the Philadelphia chromosome, a translocation mutation commonly associated with chronic myelogenous leukemia and less often with acute lymphoblastic leukemia.
Diagnostic Findings
-
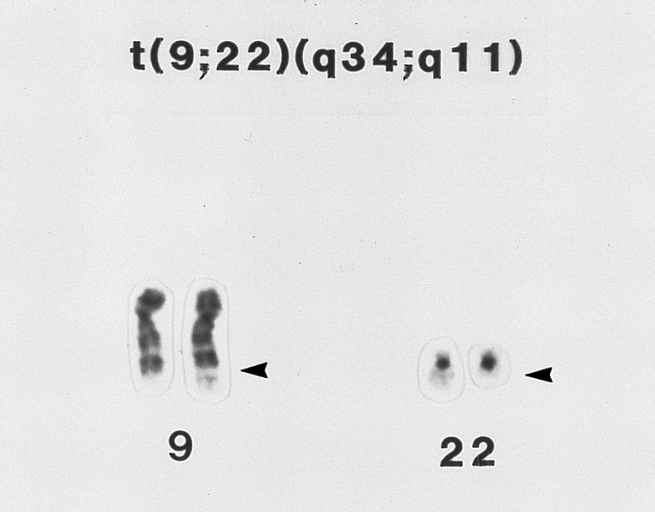
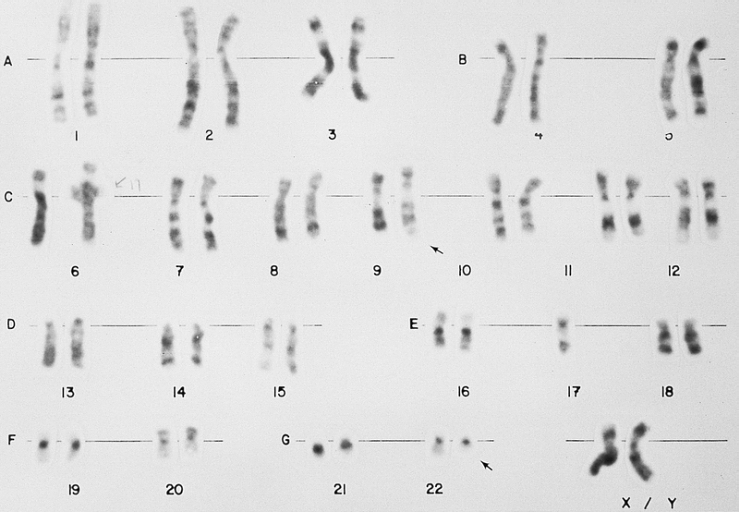 A t(9;22)(q34;q11) (Philadelphia chromosome) lymphoblast karyotype. This karyotype is from the bone marrow of a 28-year-old man with ALL. Material from the long arm (q) of chromosome 22 is translocated to the long arm of chromosome 9 (arrows). This translocation is found in approximately 20 to 25 percent of adults and approximately 2 percent of children with ALL. It is associated with a poor prognosis. (Giemsa-trypsin banding)
A t(9;22)(q34;q11) (Philadelphia chromosome) lymphoblast karyotype. This karyotype is from the bone marrow of a 28-year-old man with ALL. Material from the long arm (q) of chromosome 22 is translocated to the long arm of chromosome 9 (arrows). This translocation is found in approximately 20 to 25 percent of adults and approximately 2 percent of children with ALL. It is associated with a poor prognosis. (Giemsa-trypsin banding) -
 Acute myelogenous leukemia, AML, ANLL, M2 (FAB), karyotype; t(8;21) is specific for M2 ANLL involving possible oncogens c-mos or c-myc.
Acute myelogenous leukemia, AML, ANLL, M2 (FAB), karyotype; t(8;21) is specific for M2 ANLL involving possible oncogens c-mos or c-myc. -
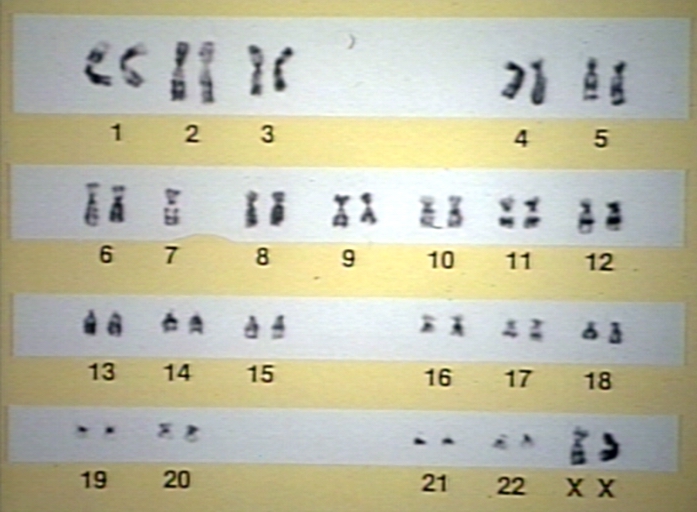 Acute myelogenous leukemia, Myelodysplastic syndrome, karyotype; Monosomy 7 or -7 is associated with bad prognosis and increased sepsis due to poor granulocyte function.
Acute myelogenous leukemia, Myelodysplastic syndrome, karyotype; Monosomy 7 or -7 is associated with bad prognosis and increased sepsis due to poor granulocyte function.
(q34;q11)_(PHILADELPHIA_CHROMOSOME)_LYMPHOBLAST_KARYOTYPE.jpg)
,_karyotype.jpg)

-
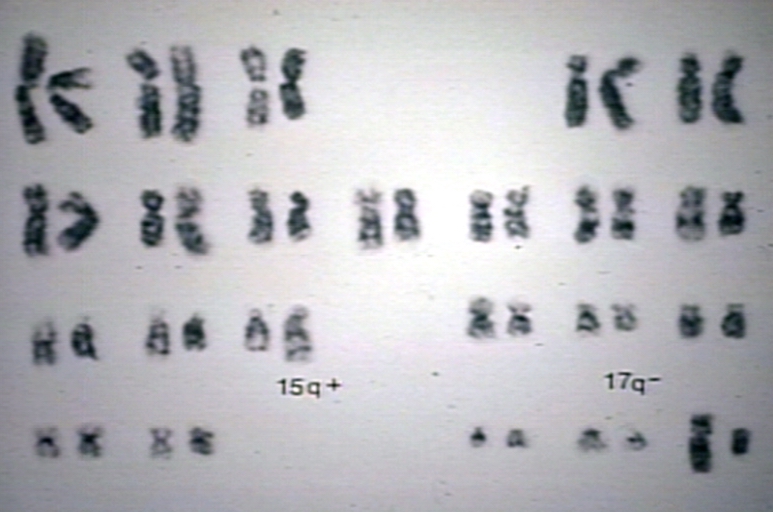 Acute Promyelocytic Leukemia, AML, M3 (FAB); t(15;17) is consistently associated with APL resulting in a fusion gene for PML and RAR-alpha which accounts for response to retinoid therapy. RAR is retinoic acid receptor.
Acute Promyelocytic Leukemia, AML, M3 (FAB); t(15;17) is consistently associated with APL resulting in a fusion gene for PML and RAR-alpha which accounts for response to retinoid therapy. RAR is retinoic acid receptor. -
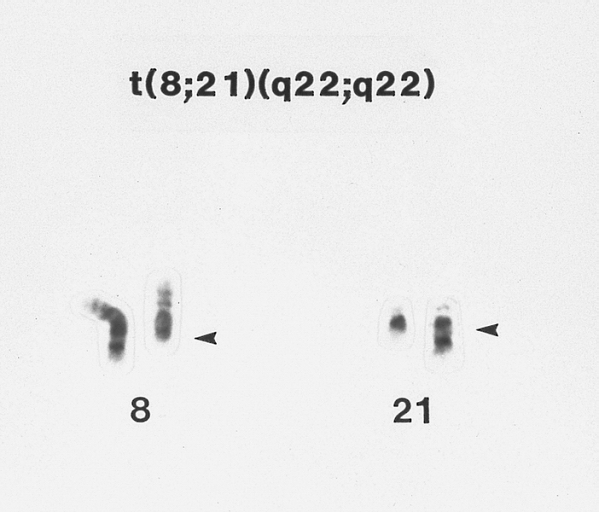 Acute myeloblastic leukemia with maturation (AML-M2) associated with A t(8;21) chromosome abnormality G-banded Wright-Giemsa stained partial karyotype showing the 8;21 chromosome translocation. The abnormal chromosomes are on the right, with breakpoints designated by arrowheads.
Acute myeloblastic leukemia with maturation (AML-M2) associated with A t(8;21) chromosome abnormality G-banded Wright-Giemsa stained partial karyotype showing the 8;21 chromosome translocation. The abnormal chromosomes are on the right, with breakpoints designated by arrowheads. -
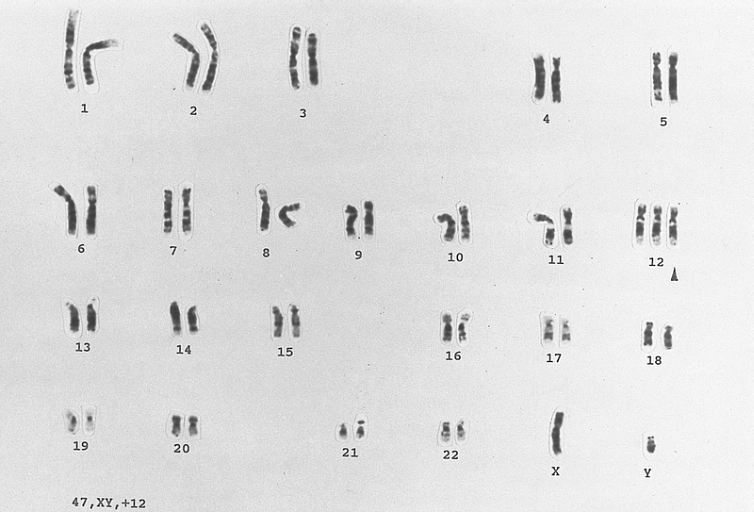 B-cell chronic lymphocytic leukemia: Trisomy for chromosome 12 Karyotype of a lymphocyte from a patient with B CLL showing an extra copy of chromosome 12. (G-banded, Wright-Giemsa stained)
B-cell chronic lymphocytic leukemia: Trisomy for chromosome 12 Karyotype of a lymphocyte from a patient with B CLL showing an extra copy of chromosome 12. (G-banded, Wright-Giemsa stained)



-
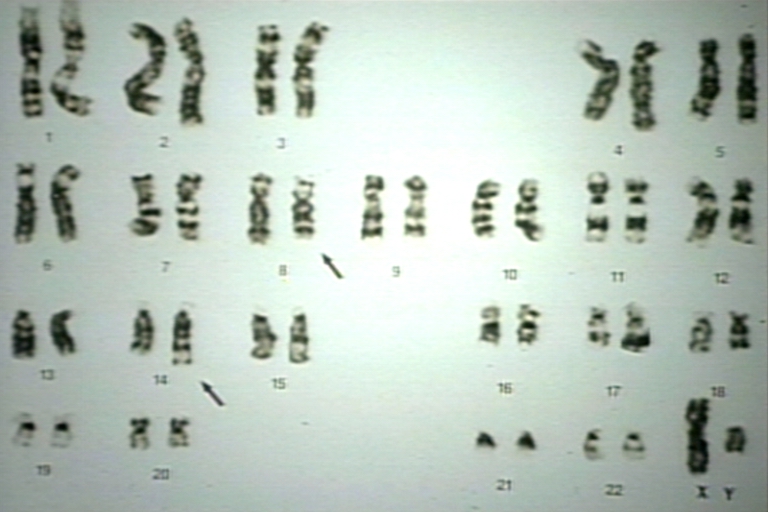 Lymph node: Burkitt's lymphoma, NonHodgkin's Lymphoma, NHL, karyotype; t(8;14) seen in Burkitt's lymphoma or L3 ALL results in a fusion protein of c-myc oncogene (8) and the Ig heavy chain gene in 14.
Lymph node: Burkitt's lymphoma, NonHodgkin's Lymphoma, NHL, karyotype; t(8;14) seen in Burkitt's lymphoma or L3 ALL results in a fusion protein of c-myc oncogene (8) and the Ig heavy chain gene in 14. -
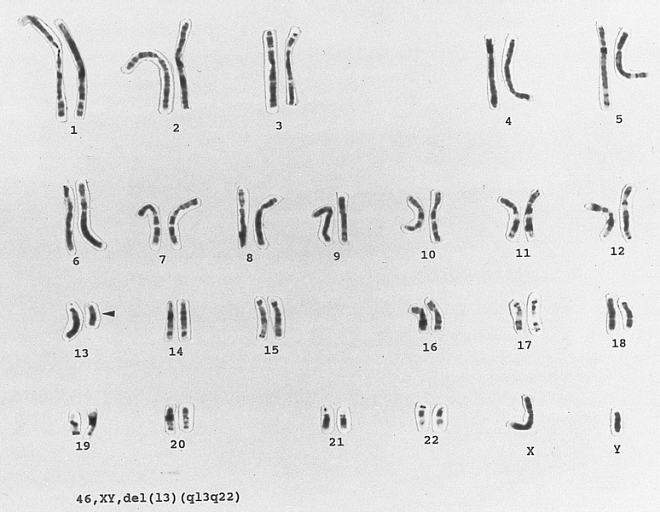 Chronic Lymphocytic Leukemia: 13q-chromosome abnormality karyotype of a lymphocyte from a patient with newly diagnosed B CLL. There is a clonal abnormality involving partial deletion of the long arm of chromosome 13 at bands q13q22. (G-banded, Wright-Giemsa stained)
Chronic Lymphocytic Leukemia: 13q-chromosome abnormality karyotype of a lymphocyte from a patient with newly diagnosed B CLL. There is a clonal abnormality involving partial deletion of the long arm of chromosome 13 at bands q13q22. (G-banded, Wright-Giemsa stained) -
 Chronic myelogenous leukemia, in transition, blast crisis, karyotype; Ph1 t(9;22), +8, i(17) are additional karyotypic changes from Ph chromosome indicating multistep in transformation to acute leukemia.
Chronic myelogenous leukemia, in transition, blast crisis, karyotype; Ph1 t(9;22), +8, i(17) are additional karyotypic changes from Ph chromosome indicating multistep in transformation to acute leukemia.



-
 Chronic Myeloid Leukemia: Philadelphia Chromosome. Partial karyotype of a myeloid cell from the bone marrow of a patient with CML showing the typical t(9;22) translocation. (G-banded, Wright-Giemsa stained)
Chronic Myeloid Leukemia: Philadelphia Chromosome. Partial karyotype of a myeloid cell from the bone marrow of a patient with CML showing the typical t(9;22) translocation. (G-banded, Wright-Giemsa stained) -
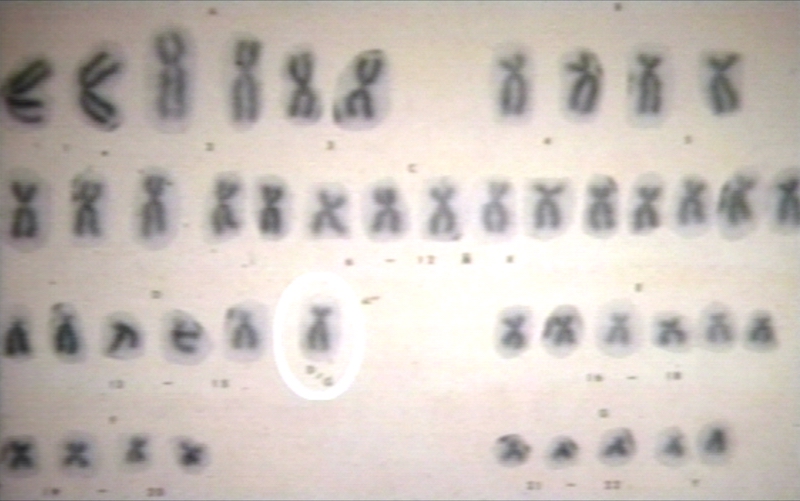 Down syndrome G karyotype
Down syndrome G karyotype -
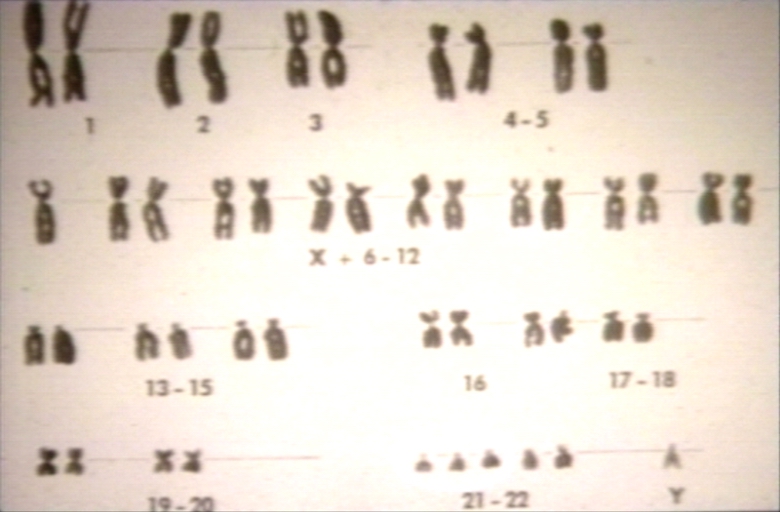 Down syndrome karyotype
Down syndrome karyotype



-
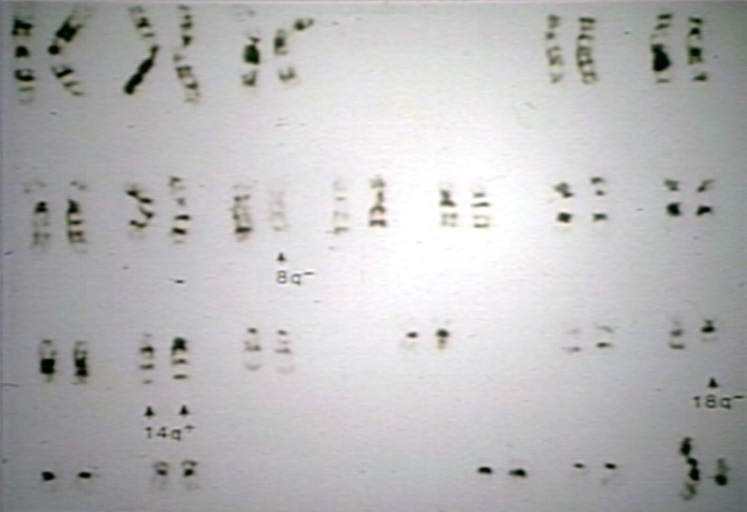 Lymph node: Evolved B cell NonHodgkin's lymphoma, NHL; t(8;14) and t(14;18), appears in 75% of follicular NHL, juxtaposing the bcl-2 (band 18q21) proto-oncogene with Ig (band 14q32) heavy chain gene (resulting in deregulation of bcl-2 (inhibits apoptosis)
Lymph node: Evolved B cell NonHodgkin's lymphoma, NHL; t(8;14) and t(14;18), appears in 75% of follicular NHL, juxtaposing the bcl-2 (band 18q21) proto-oncogene with Ig (band 14q32) heavy chain gene (resulting in deregulation of bcl-2 (inhibits apoptosis) -
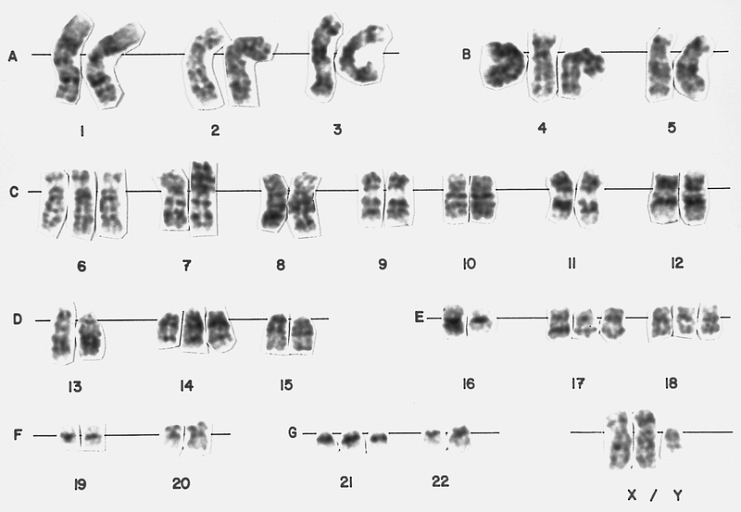 Hyperdiploid lymphoblast karyotype This bone marrow karyotype is from a 5-year-old female with ALL. There is hyperdiploidy with 55 chromosomes. Hyperdiploidy with greater than 50 chromosomes is found in approximately 25 percent of children with ALL and is associated with a good prognosis. (Giemsa-trypsin banding)
Hyperdiploid lymphoblast karyotype This bone marrow karyotype is from a 5-year-old female with ALL. There is hyperdiploidy with 55 chromosomes. Hyperdiploidy with greater than 50 chromosomes is found in approximately 25 percent of children with ALL and is associated with a good prognosis. (Giemsa-trypsin banding) -
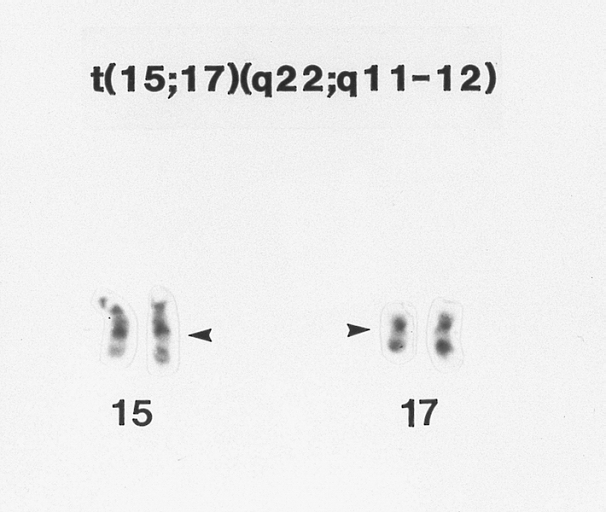 Hypergranular acute promyelocytic leukemia (AML-M3) G-banded Wright-Giemsa stained partial karyotype showing the 15;17 chromosome translocation. The breakpoints on the abnormal chromosomes are designated by arrowheads.
Hypergranular acute promyelocytic leukemia (AML-M3) G-banded Wright-Giemsa stained partial karyotype showing the 15;17 chromosome translocation. The breakpoints on the abnormal chromosomes are designated by arrowheads.



-
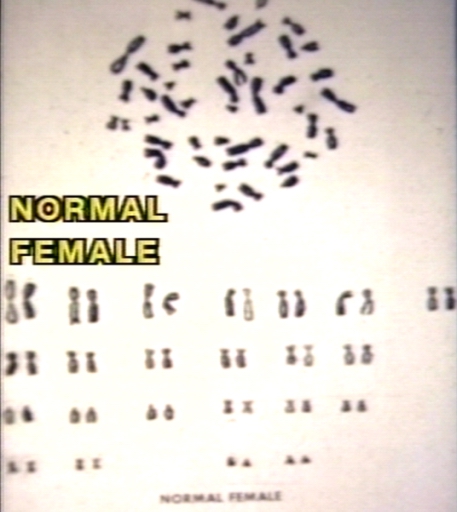
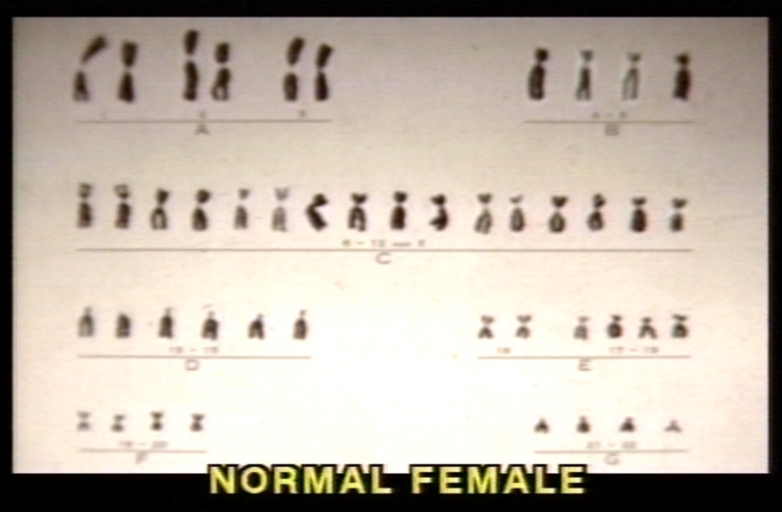 Karyotype of normal female.
Karyotype of normal female. -
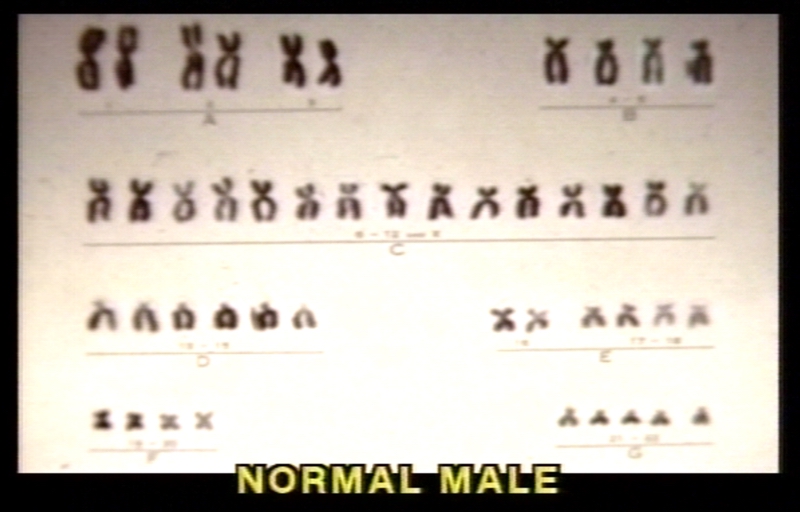 Karyotype of normal male.
Karyotype of normal male. -
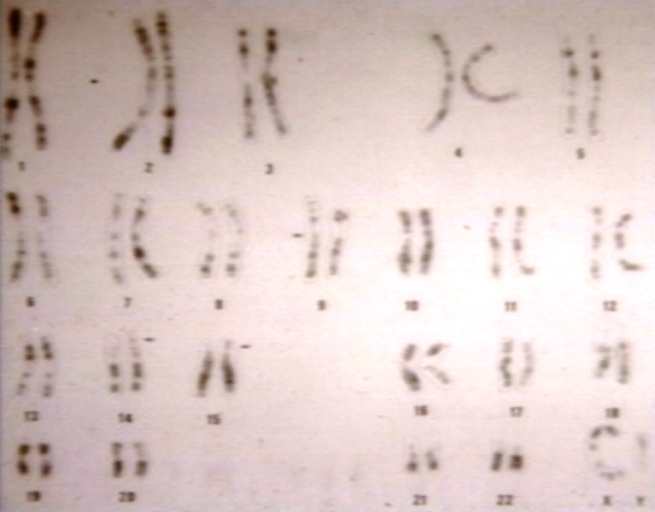 Karyotype R bands composite
Karyotype R bands composite



-
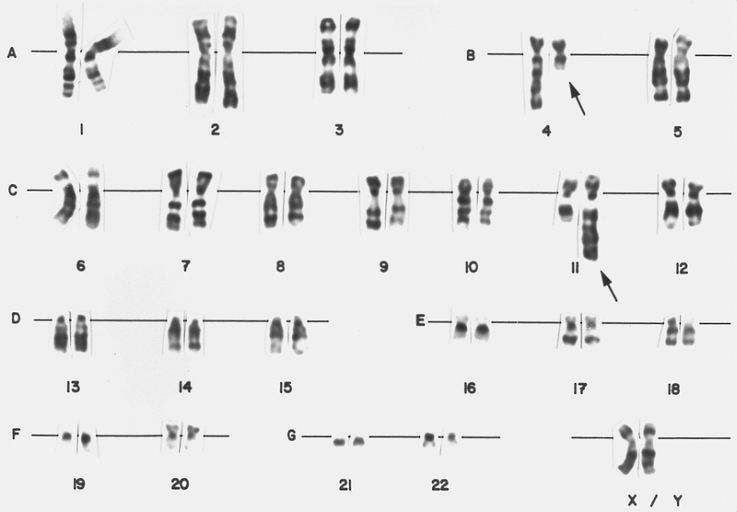 Lymphoblast karyotype from a patient with mixed phenotype acute leukemia (Bilineal). A karyotype from bone marrow cells of the patient whose blood smear is depicted in figure 146. There is a translocation of material from the long arm of chromosome 4 to the long arm of chromosome 11, t(4;11) (q21;q23). This translocation is found in less than 5 percent of cases of childhood ALL. It is associated with markedly elevated leukocyte counts and poor prognosis. Approximately 20 percent of patients are less than 1 month of age at diagnosis (congenital leukemia).
Lymphoblast karyotype from a patient with mixed phenotype acute leukemia (Bilineal). A karyotype from bone marrow cells of the patient whose blood smear is depicted in figure 146. There is a translocation of material from the long arm of chromosome 4 to the long arm of chromosome 11, t(4;11) (q21;q23). This translocation is found in less than 5 percent of cases of childhood ALL. It is associated with markedly elevated leukocyte counts and poor prognosis. Approximately 20 percent of patients are less than 1 month of age at diagnosis (congenital leukemia). -
 Myelodysplastic syndrome (MDS), Agnogenic myeloid metaplasia, myelofibrosis, karyotype; Trisomy 8 or +8 is most common karyotype abnormality in MDS, and acute nonlymphocytic leukemias.
Myelodysplastic syndrome (MDS), Agnogenic myeloid metaplasia, myelofibrosis, karyotype; Trisomy 8 or +8 is most common karyotype abnormality in MDS, and acute nonlymphocytic leukemias. -
 Myelodysplastic syndrome, MDS. Refractory anemia (RA), karyotype, 5q- syndrome; 5q- abnormality per se is not a bad prognostic marker but results in a refractory anemia called 5q- syndrome.
Myelodysplastic syndrome, MDS. Refractory anemia (RA), karyotype, 5q- syndrome; 5q- abnormality per se is not a bad prognostic marker but results in a refractory anemia called 5q- syndrome.
.jpg)
,_Agnogenic_myeloid_metaplasia,_myelofibrosis,_karyotype.jpg)
,_karyotype.jpg)
-
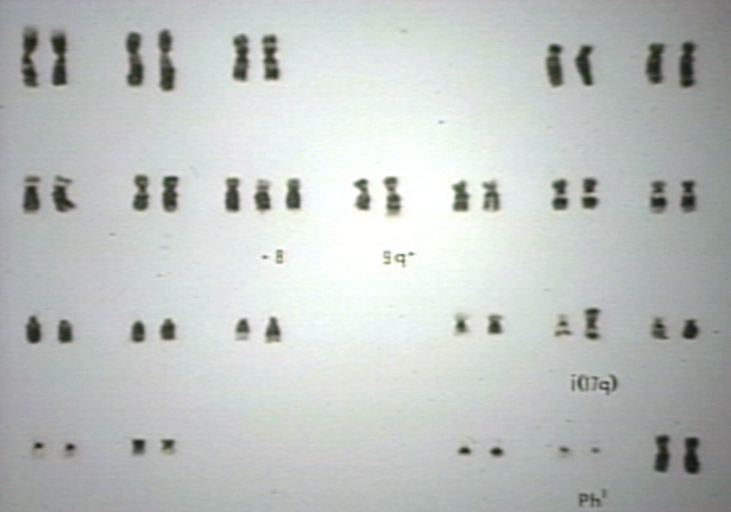 Myelodysplastic syndrome, MDS. Refractory anemia (RA), karyotype, 5q- syndrome; 5q- abnormality per se is not a bad prognostic marker but results in a refractory anemia called 5q- syndrome.
Myelodysplastic syndrome, MDS. Refractory anemia (RA), karyotype, 5q- syndrome; 5q- abnormality per se is not a bad prognostic marker but results in a refractory anemia called 5q- syndrome. -
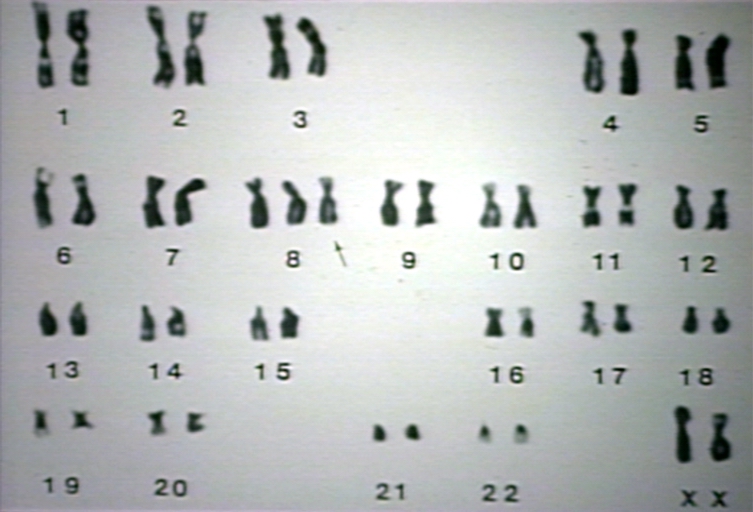
 Normal female karyotype.
Normal female karyotype. -
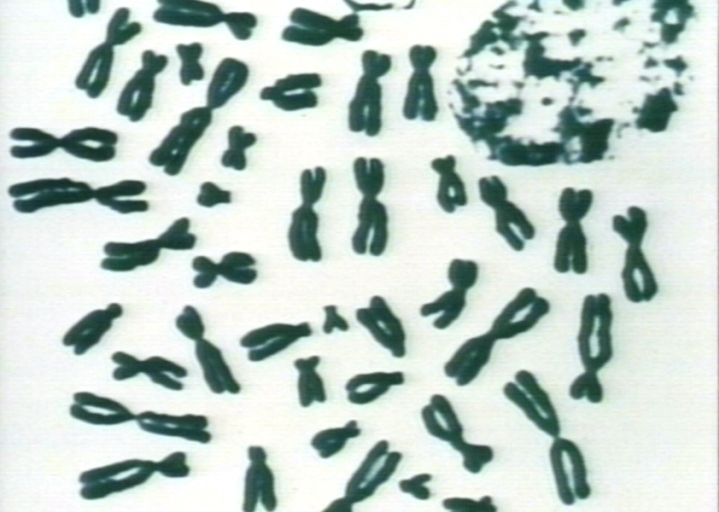 Normal karyotype.
Normal karyotype.



-
 Philadelphia chromosome in Chronic Myelogenous Leukemia (CML); Ph, t(9;22) results in a fusion mRNA of 210 kd called bcr/abl which inhibits cellular apoptosis and increase granulocytes in Chronic Myelogenous Leukemia (CML).
Philadelphia chromosome in Chronic Myelogenous Leukemia (CML); Ph, t(9;22) results in a fusion mRNA of 210 kd called bcr/abl which inhibits cellular apoptosis and increase granulocytes in Chronic Myelogenous Leukemia (CML). -
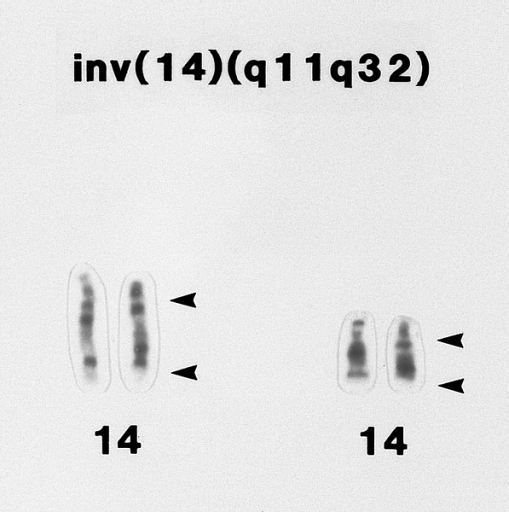 T-prolymphocytic leukemia. Portion of a karyotype of a lymphocyte from the specimen shown in B showing an inv(14)(q11q32) abnormality. (G banded, Wright-Giemsa stained)
T-prolymphocytic leukemia. Portion of a karyotype of a lymphocyte from the specimen shown in B showing an inv(14)(q11q32) abnormality. (G banded, Wright-Giemsa stained) -
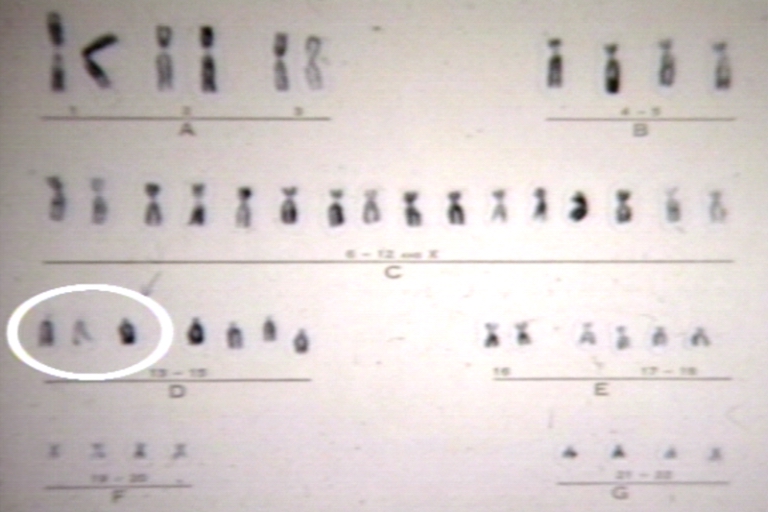 Trisomy 13 karyotype.jpg
Trisomy 13 karyotype.jpg



-
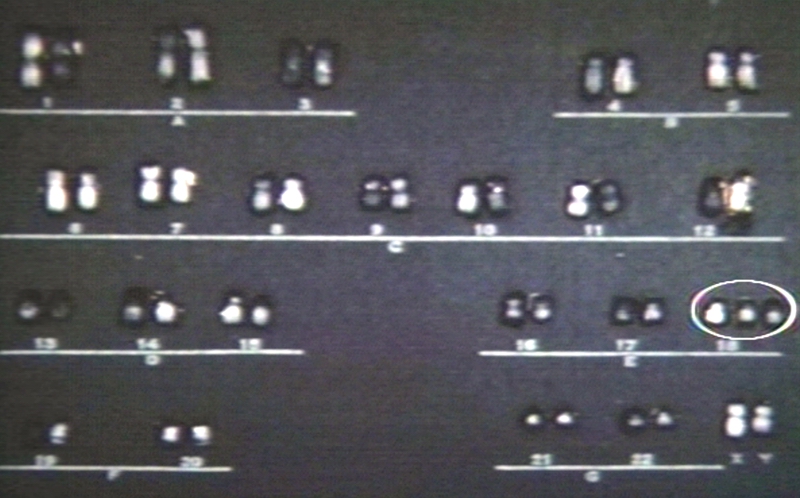 Trisomy 18 karyotype.jpg
Trisomy 18 karyotype.jpg -
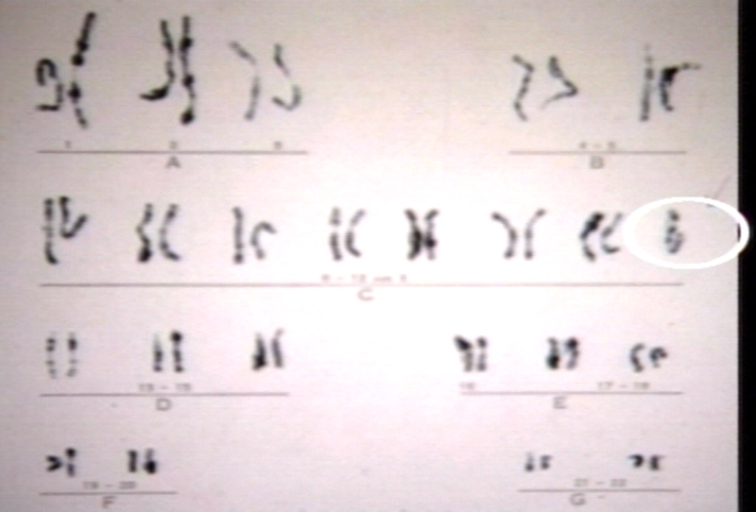 Turner's karyotype.
Turner's karyotype. -
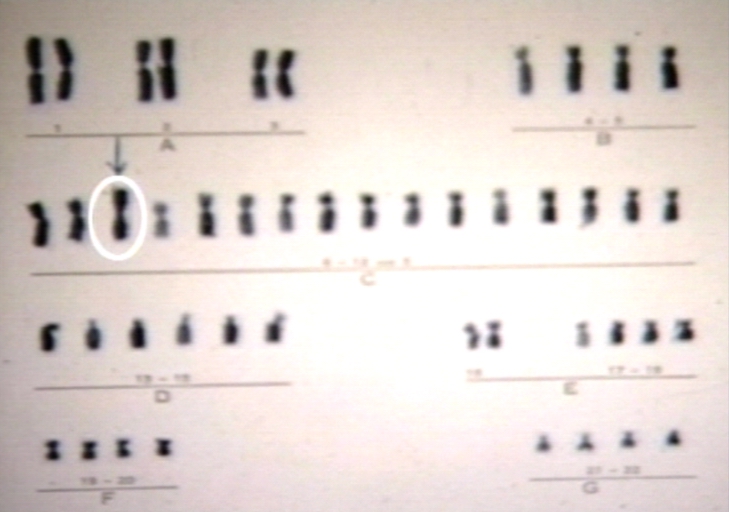 Turner's karyotype 46x 150x.
Turner's karyotype 46x 150x.



See also
References
- ↑ White M.J.D. 1973. The chromosomes. 6th ed, Chapman & Hall, London. p28
- ↑ Stebbins G.L. 1950. Variation and evolution in plants. Chapter XII: The Karyotype. Columbia University Press N.Y.
- ↑ Gustashaw K.M. 1991. Chromosome stains. In The ACT Cytogenetics Laboratory Manual 2nd ed, ed. M.J. Barch. The Association of Cytogenetic Technologists, Raven Press, New York.
- ↑ Stebbins G.L. 1971. Chromosomal evolution in higher plants. Arnold, London. p85-6
- ↑ White M.J.D. 1973. Animal cytology and evolution. 3rd ed, Cambridge University Press.
- ↑ Levitsky G.A. 1924. The material basis of heredity. State Publication Office of the Ukraine, Kiev. [in Russian]
- ↑ Levitsky G.A. 1931. The morphology of chromosomes. Bull. Applied Bot. Genet. Plant Breed. 27, 19-174.
- ↑ Darlington C.D. 1939. Evolution of genetic systems. Cambridge University Press. 2nd ed, revised and enlarged, 1958. Oliver & Boyd, Edinburgh.
- ↑ White M.J.D. 1973. Animal cytology and evolution. 3rd ed, Cambridge University Press.
- ↑ White M.J.D. 1973. The chromosomes. 6th ed, Chapman & Hall, London.
- ↑ Kottler M. 1974. From 48 to 46: cytological technique, preconception and the counting of the human chromosomes. Bull. Hist. Med. 48, 465-502.
- ↑ von Winiwarter H. 1912. Études sur la spermatogenese humaine. Arch. biologie 27, 93, 147-9.
- ↑ Painter T.S. 1922. The spermatogenesis of man. Anat. Res. 23, 129.
- ↑ Painter T.S. 1923. Studies in mammalian spermatogenesis II. The spermatogenesis of man. J. Exp. Zoology 37, 291-336.
- ↑ Tjio J.H & Levan A. 1956. The chromosome number of man. Hereditas 42, 1-6.
- ↑ Hsu T.C. Human and mammalian cytogenetics: a historical perspective. Springer-Verlag, N.Y.
- ↑ Goday C. and Esteban M.R. 2001. Chromosome elimination in sciarid flies. Bioessays 23: 242–250.
- ↑ Müller F. Bernard V. & Tobler H. 1996. Chromatin diminution in nematodes. Bioessays 18: 133–138.
- ↑ Wyngaard G.A. & Gregory T.R. 2001. Temporal control of DNA replication and the adaptive value of chromatin diminution in copepods. J. Exp. Zool. 291: 310–16.
- ↑ Gilbert S.F. 2006. Developmental biology. Sinauer Associates, Stamford CT. 8th ed, Chapter 9
- ↑ Wurster D.H. and Benirschke K. 1970. Indian Muntjac, Muntiacus muntjak: a deer with a low diploid number. Science 168, 1364-1366.
- ↑ Hsu T.C. 1979. Human and mammalian cytogenetics: an historical perspective. Springer-Verlag N.Y. p73-4
- ↑ Khandelwal S. 1990. Chromosome evolution in the genus Ophioglossum L. Botanical Journal of the Linnean Society 102: 205–217.
- ↑ Kim, D.S. (2005). "Karyotype of North American shortnose sturgeon Acipenser brevirostrum with the highest chromosome number in the Acipenseriformes" (PDF). Ichthyological Research. 52 (1): 94–97. Retrieved 2008-03-18. Unknown parameter
|coauthors=ignored (help) - ↑ Stebbins, G.L. 1940. The significance of polyploidy in plant evolution. The American Naturalist 74, 54–66.
- ↑ Stebbins G.L. 1950. Variation and evolution in plants. Columbia University Press.
- ↑ Comai L. 2005. The advantages and disadvantages of being polyploid. Nature Reviews, Genetics. 6, 836-46.
- ↑ Adams K.L. & Wendel J.F. 2005. Polyploidy and genome evolution in plants. Current Opinion in Plant Biology. 8 135-41.
- ↑ Stebbins G.L. 1970. Chromosomal evolution in flowering plants. Arnold, London.
- ↑ Gregory T.R. & Mable B.K. 2005. Polyploidy in animals. In The Evolution of the genome Gregory T.R. (ed). Elsevier, San Diego. p427-517
- ↑ White M.J.D. 1973. The chromosomes. 6th ed, Chapman & Hall, London. p45
- ↑ Lilly M.A. & Duronio R.J. (2005). "New insights into cell cycle control from the Drosophila endocycle". Oncogene. 24 (17): 2765–75. PMID 15838513.
- ↑ Edgar B.A. & Orr-Weaver T.L. 2001. Endoreduplication cell cycles: more for less. Cell 105, 297-306.
- ↑ Nagl W. 1978. Endopolyploidy and polyteny in differentiation and evolution: towards an understanding of quantitative and qualitative variation of nuclear DNA in ontogeny and phylogeny. Elsevier, New York.
- ↑ Clague D.A. & G.B. Dalrymple. 1987. The Hawaiian-Emperor volcanic chain, Part I. Geologic evolution. In R.W. Decker, T.L. Wright & P.H. Stauffer (eds) Volcanism in Hawaii. U.S. Geological Survey Professional Paper 1350, 5-54.
- ↑ Carson H.L. 1970. Chromosomal tracers of evolution. Science 168, 1414–1418.
- ↑ Carson H.L. 1983. Chromosomal sequences and interisland colonizations in Hawaiian Drosophila. Genetics 103, 465-482.
- ↑ Carson H.L. 1992. Inversions in Hawaiian Drosophila. In: Krimbas C.B. & Powell J.R. (eds) Drosophila inversion polymorphism. CRC Press, Boca Raton, FL. 407-439.
- ↑ Kaneshiro K.Y. Gillespie R.G. and Carson H.L. 1995. Chromosomes and male genitalia of Hawaiian Drosophila: tools for interpreting phylogeny and geography. In Wagner W.L. & Funk E. (eds) Hawaiian biogeography: evolution on a hot spot archipelago, Smithsonian Institute Press, Washington D.C. 57-71
- ↑ Craddock E.M. 2000. Speciation processes in the adaptive radiation of Hawaiian plants and animals. Evolutionary Biology 31, 1-43.
- ↑ Ziegler A.C. 2002. Hawaiian natural history, ecology and evolution. Honolulu: University of Hawaii Press.
- ↑ Maynard Smith J. 1998. Evolutionary genetics. 2nd ed, Oxford. p218-9
- ↑ Gustashaw K.M. 1991. Chromosome stains. In The ACT Cytogenetics Laboratory Manual 2nd ed, ed. M.J. Barch. The Association of Cytogenetic Technologists, Raven Press, New York.
- ↑ Lisa G. Shaffer, Niels Tommerup (ed.). ISCN 2005: An International System for Human Cytogenetic Nomenclature. Switzerland: S. Karger AG. ISBN 3-8055-8019-3.
- ↑ E. Schröck, S. du Manoir, T. Veldman, B. Schoell, J. Wienberg, M. A. Ferguson-Smith, Y. Ning, D. H. Ledbetter, I. Bar-Am, D. Soenksen, Y. Garini, T. Ried. Multicolor spectral karyotyping of human chromosomes. Science, 26 July 1996; 273 (5274):494. abstract
- ↑ Digital karyotyping - Wang et al., 10.1073/pnas.202610899 - Proceedings of the National Academy of Sciences
cs:Karyotyp de:Karyogramm fa:کاریوتیپ it:Cariotipo he:קריוטיפ lt:Kariotipas nl:Karyotype no:Karyotype sr:Кариотип fi:Karyotyyppi sv:Karyotyp