Cochlea
Template:Infobox Anatomy The cochlea is the auditory portion of the inner ear. Its core component is the Organ of Corti, the sensory organ of hearing, which is distributed along the partition separating fluid chambers in the coiled tapered tube of the cochlea.
The name is from the Latin for snail, which is from the Greek kokhlias "snail, screw," from kokhlos "spiral shell,"(etymology) in reference to its coiled shape; the cochlea is coiled in most mammals, monotremes being the exceptions.
Anatomy

Structures
The cochlea is a spiralled, hollow, conical chamber of bone. Its structures include:
- the scala vestibuli (containing perilymph), which lies superior to the cochlear duct and abuts the oval window.
- the scala tympani (containing perilymph), which lies inferior to the scala media and terminates at the round window.
- the scala media (containing endolymph), which is the membraneous cochlear duct containing the organ of Corti.
- the helicotrema is the location where the scala tympani and the scala vestibuli merge
- Reissner's membrane separates the scala vestibuli from the scala media.
- The basilar membrane, a main structural element that determines the mechanical wave propagation properties of the cochlear partition, separates the scala media from the scala tympani.
- The Organ of Corti is the sensory epithelium, a cellular layer on the basilar membrane, powered by the potential difference between the perilymph and the endolymph. It is lined with hair cells—sensory cells topped with hair-like structures called stereocilia.
Function
In brief: the cochlea is filled with a watery liquid, which moves in response to the vibrations coming from the middle ear via the oval window. As the fluid moves, thousands of "hair cells" are set in motion, and convert that motion to electrical signals that are communicated via neurotransmitters to many thousands of nerve cells. These primary auditory neurons transform the signals into electrical impulses known as action potentials, which travel along the auditory nerve to structures in the brainstem for further processing.
The stapes of the middle ear transmits to the fenestra ovalis (oval window) on the outside of the cochlea, which vibrates the perilymph (fluid) in the scala vestibuli (upper chamber of the cochlea).
This motion of perilymph in turn vibrates the endolymph in the scala media, the perilymph in the scala tympani, the basilar membrane, and organ of Corti, thus causing movements of the hair bundles of the hair cells, acoustic sensor cells that convert vibration into electrical potentials. The hair cells in the organ of Corti are tuned to certain sound frequencies[1], being responsive to high frequencies near the oval window and to low frequencies near the apex of the cochlea.
The hair cells are arranged in four rows in the organ of Corti along the entire length of the cochlear coil. Three rows consist of outer hair cells (OHCs) and one row consists of inner hair cells (IHCs). The inner hair cells provide the main neural output of the cochlea. The outer hair cells, instead, mainly receive neural input from the brain, which influences their motility as part of the cochlea’s mechanical pre-amplifier. The input to the OHC is from the olivary body via the medial olivocochlear bundle.
For very low frequencies (below 20Hz), the pressure waves propagate along the complete route of the cochlea - up scala vestibuli, around helicotrema and down scala tympani to the round window. Frequencies this low do not activate the organ of Corti and are below the threshold for hearing. Higher frequencies do not propagate to the helicotrema but are transmitted through the endolymph in the cochlear duct to the perilymph in the scala tympani.
A very strong movement of the endolymph due to very loud noise may cause hair cells to die. This is a common cause of partial hearing loss and is the reason why users of firearms or heavy machinery should wear earmuffs or earplugs.
Detailed anatomy
The walls of the hollow cochlea are made of bone, with a thin, delicate lining of epithelial tissue. This coiled tube is divided through most of its length by a membrane partition. Two fluid-filled spaces (scalae) are formed by this dividing membrane.
The fluid in both is called perilymph: a clear solution of electrolytes and proteins. The two scalae (fluid-filled chambers) communicate with each other through an opening at the top (apex) of the cochlea called the helicotrema, a common space that is the one part of the cochlea that lacks the lengthwise dividing membrane.
At the base of the cochlea each scala ends in a membrane that faces the middle ear cavity. The scala vestibuli ends at the oval window, where the footplate of the stapes sits. The footplate rocks when the ear drum moves the ossicular chain; sending the perilymph rippling with the motion, the waves moving away from footplate and towards helicotrema. Those fluid waves then continue in the perilymph of the scala tympani. The scala tympani ends at the round window, which bulges out when the waves reach it -providing pressure relief. This one-way movement of waves from oval window to round window occurs because the middle ear directs sound to the oval window, but shields the round window from being struck by sound waves from the external ear. It is important, because waves coming from both directions, from the round and oval window would cancel each other out. In fact, when the middle ear is damaged such that there is no tympanic membrane or ossicular chain, and the round window is oriented outward rather than set under a bit of a ledge in the round window niche, there is a maximal conductive hearing loss of about 60 dB.
The lengthwise partition that divides most of the cochlea is itself a fluid-filled tube, the third scalae. This central column is called the scala media or cochlear duct. Its fluid, endolymph, also contains electrolytes and proteins, but is chemically quite different from perilymph. Whereas the perilymph is rich in sodium salts, the endolymph is rich in potassium salts.
The cochlear duct is supported on three sides by a rich bed of capillaries and secretory cells (the stria vascularis), a layer of simple squamous epithelial cells (Reissner's membrane), and the basilar membrane, on which rests the receptor organ for hearing - the organ of Corti. The cochlear duct is almost as complex on its own as the ear itself.
The ear is a very active organ. Not only does the cochlea "receive" sound, it generates it! Some of the hair cells of the cochlear duct can change their shape enough to move the basilar membrane and produce sound. This process is important in fine tuning the ability of the cochlea to accurately detect differences in incoming acoustic information. The sound produced by the inner ear is called an otoacoustic emission (OAE), and can be recorded by a microphone in the ear canal. Otoacoustic emissions are important is some types of tests for hearing impairment.
Comparative physiology
The coiled form of cochlea is unique to mammals. In birds and in other non-mammalian vertebrates the compartment containing the sensory cells for hearing is occasionally also called “cochlea”, although it is not coiled up. Instead it forms a blind-ended tube, also called the cochlear duct. This difference apparently evolved in parallel with the differences in frequency range of hearing and in frequency resolution between mammals and non-mammalian vertebrates. Most bird species do not hear above 4–5 kHz, the currently known maximum being ~ 11 kHz in the barn owl. Some marine mammals hear up to 200 kHz. The superior frequency resolution in mammals is due to their unique mechanism of pre-amplification of sound by active cell-body vibrations of outer hair cells. A long coiled compartment, rather than a short and straight one, provides more space for frequency dispersion and is therefore better adapted to the highly derived functions in mammalian hearing.[2]
As the study of the cochlea should fundamentally be focused upon the level of hair cells, it is important to note the anatomical and physiological differences between the hair cells of various species. In birds, for instance, instead of outer and inner hair cells, there are tall and short hair cells. There are several similarities of note in regard to this comparative data. For one, the tall hair cell is very similar in function to that of the inner hair cell and the short hair cell is very similar in function to that of the outer hair cell. One unavoidable difference, however, is that while all hair cells are attached to a tectorial membrane in birds, only the outer hair cells are attached to the tectorial membrane in mammals.
References
- ↑ Tasaki I. Nerve impulses in individual auditory nerve fibers of guinea pig. J Neurophysiol. 17(2): 97-122, 1954
- ↑ Vater M, Meng J, Fox RC. Hearing organ evolution and specialization: Early and later mammals. In: GA Manley, AN Popper, RR Fay (Eds). Evolution of the Vertebrate Auditory System, Springer-Verlag, New York 2004, pp 256–288.
See also
Additional images
-
Vestibular system
-
Ear labyrinth
-
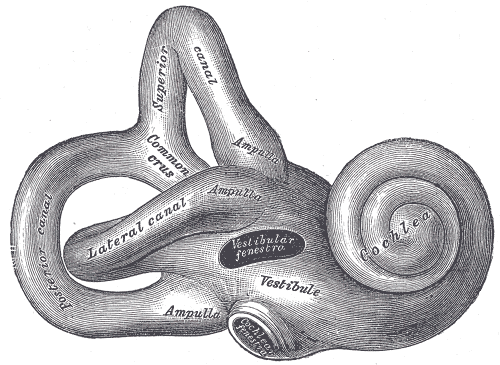
Right osseous labyrinth. Lateral view.
-
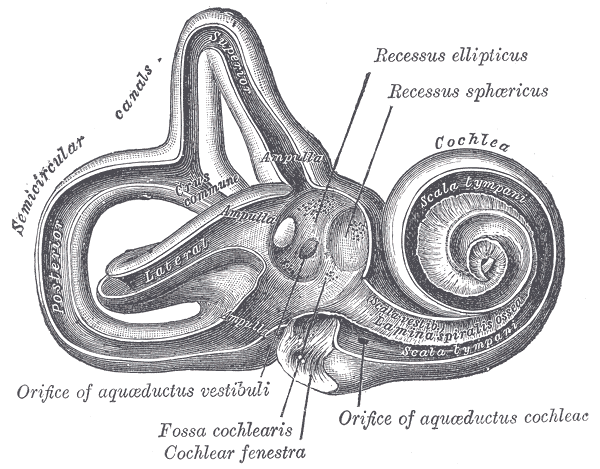
Interior of right osseous labyrinth.
-
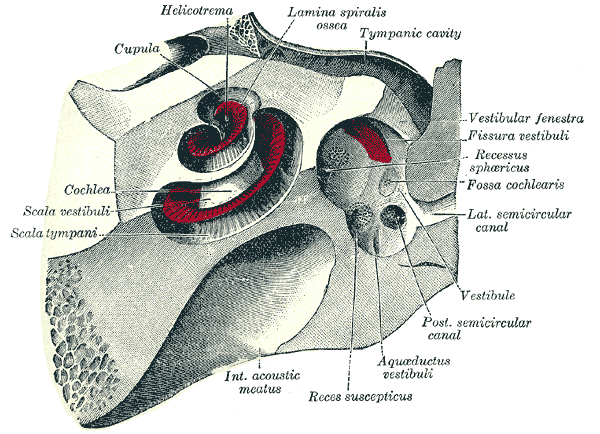
The cochlea and vestibule, viewed from above.
-
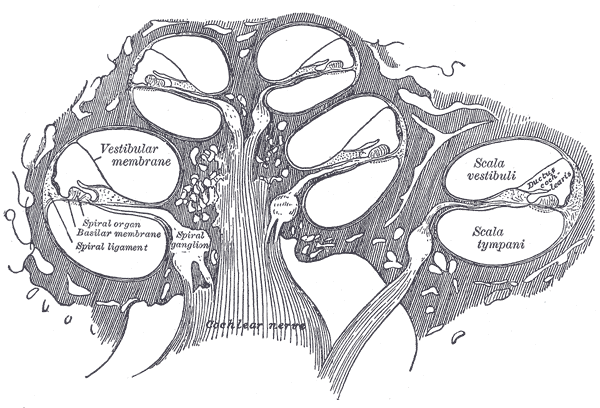
Diagrammatic longitudinal section of the cochlea.




{kind=link}
{kind=link}
External links
- Cochlea at the US National Library of Medicine Medical Subject Headings (MeSH)
- "Promenade 'Round the Cochlea" by R. Pujol, S. Blatrix S. et al. at University of Montpellier
- "The Cochlea Homepage", F. Mammano and R. Nobili of the Venetian Institute of Molecular Medicine
- MYSTERIES OF THE COCHLEA Dr. Janez Faganel Memorial Lecture at Klinični center Ljubljana
de:Innenohr hu:Csiga (cochlea) nl:Slakkenhuis (oor) sk:Slimák (kochlea) sv:Hörselsnäckan